Weevil News
|
No. 25 |
9 pp. |
10th January 2005 |
ISSN 1615-3472
|
|
|
Stüben P.E. (2005): Zur
Verbreitung von Kyklioacalles navieresi
(Boheman 1837) und Kyklioacalles roboris (Curtis 1834) im
Rheinland / Germany unter besonderer Berücksichtigung der Fundumstände auf
dem Bausenberg (Eifel). (Coleoptera: Curculionidae: Cryptorhynchinae) - Weevil News: http://www.curci.de/Inhalt.html, No. 25: 9 pp., CURCULIO-Institute: Mönchengladbach. (ISSN 1615-3472). |
|||||
Zur Verbreitung von Kyklioacalles
navieresi (Boheman 1837) und Kyklioacalles roboris (Curtis 1834) im Rheinland / Germany unter besonderer
Berücksichtigung der Fundumstände auf dem Bausenberg (Eifel)
(Coleoptera: Curculionidae: Cryptorhynchinae)
von
Peter E. Stüben, Mönchengladbach*
mit
28 Abbildungen
[Stüben]
Abstract
Distribution of Kyklioacalles navieresi (Boheman, 1837) and Kyklioacalles roboris (Curtis, 1834) in the Rhineland / Germany and
finding circumstances at the Bausenberg / Eifel Mountains (Coleoptera: Curculionidae:
Cryptorhynchinae); with 28 figures.
Starting from differential diagnosis, identification of food and host
plants and of Palaearctic and parapatric distribution, the occurrence of the
species Kyklioacalles navieresi
(Boheman, 1837) and Kyklioacalles roboris
(Curtis, 1834) in the Rhineland is presented and discussed. Both species were
found at the Bausenberg / Eifel Mountains for the first time in the same area.
The hypothesis, that Kyklioacalles
roboris is the more hygrophilous
species over large parts of Europe, must be completed by the hypothesis that Kyklioacalles navieresi clearly prefers more xerothermous sites. On
the southern sun-exposed slopes of the Bausenberg, a former volcano of Eifel
Mountains, with a sparse and dry vegetation of Quercus robur shrubs,
exclusively Kyklioacalles
navieresi could be found. In the
summer month, this part of the Bausenberg is often drying out and heating up
within short periods. Based on experiences with other Kyklioacalles species, the two first larval stages of Kyklioacalles roboris
cannot develop in such dry habitats; they need three month in high summer.
However, they have a better chance in dense deciduous forests or in hedgerows
with Carpinus betulus L. Moist and shady habitats are clearly
preferred by Kyklioacalles roboris
at the Bausenberg.
Key Words
Coleoptera,
Curculionidae, Cryptorhynchinae, Kyklioacalles
navieresi, Kyklioacalles roboris,
biology, ecology, host-plant, Germany, Rhineland,
Bausenberg.
Ausgehend von
der Differentialdiagnose, der Darstellung der Fraß-
und Entwicklungspflanzen sowie der paläarktischen und der parapatrischen
Verbreitung der Arten Kyklioacalles
navieresi (Boheman 1837) und Kyklioacalles roboris (Curtis 1834) wird das Vorkommen im Rheinland, insbesondere
das auf dem Bausenberg (Eifel) erstmalig beobachtete gemeinsame Auftreten beider Arten, dargestellt und diskutiert.
Die These, dass in weiten Teilen
Europas Kyklioacalles
roboris die hygrophilere
Art ist, muss um die These, dass Kyklioacalles navieresi die eindeutig
xerothermere Standorte präferierende Art ist, ergänzt werden. So findet sich
auf den Südhängen des Bausenbergs, eines ehemaligen Eifel-Vulkans, in den
lichten, trockenen Quercus robur -
Gebüschen ausschließlich Kyklioacalles navieresi ! Dieser Teil des Bausenbergs kann rasch austrocknen und sich
unter der offenen Vegetation in den Sommermonaten sehr stark erwärmen. Legt man
die Erfahrungen mit anderen Kyklioacalles-Arten
zugrunde, werden sich die Larven von Kyklioacalles
roboris - während der dreimonatigen Entwicklung im Hochsommer - in den
ersten beiden Larvalstadien mit solchen trockenen Habitatstrukturen nicht
abfinden, hier also keine geeigneten Entwicklungsmöglichkeiten vorfinden.
Hingegen werden sie diese Entwicklungsmöglichkeiten eher in den dichteren
Laubwäldern oder unter Hecken (z.B. an Carpinus
betulus L.) antreffen. Es sind die feuchten und schattigen
Habitatstrukturen, die die Imagines von Kyklioacalles
roboris auch auf dem Bausenberg eindeutig präferieren.
In meiner "Revision des Genus Kyklioacalles und der Beschreibung der Untergattung Palaeoacalles subg. n. unter
Heranziehung phylogenetischer, morphogenetischer und biogeographischer
Aspekte" aus dem Jahre 2003 habe ich die Art Acalles navieresi Boheman 1837 resynonymisiert und
zusammen mit der Art Acalles roboris Curtis
1834 in die Gattung Kyklioacalles Stüben 1999 transferiert und
redeskribiert. [Stüben 2003a: 116-166]
Das neue Subgenus Palaeoacalles ist bis auf wenige Ausnahmen - den hohen Norden Europas
und den nordafrikanischen Raum - in der ganzen Westpaläarktis (mit dem
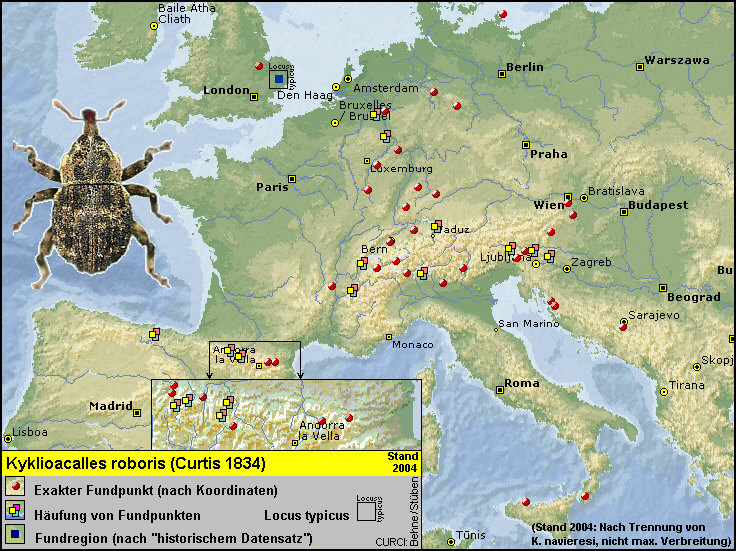
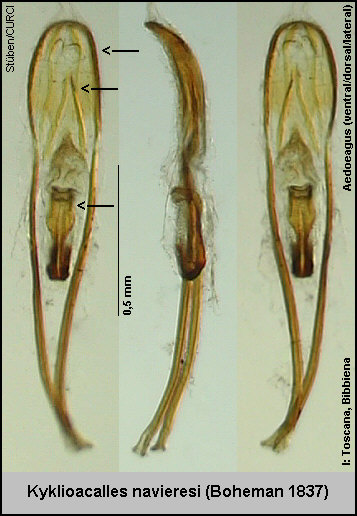
Schwerpunkt in Mitteleuropa) verbreitet und wird hier von den drei Arten Kyklioacalles roboris [Fig. W25.1], Kyklioacalles
navieresi
[Fig. W25.2] und einer noch
neu zu beschreibenden Art aus Kroatien [Stüben 2005e] vertreten.
Es ist - so scheint es zunächst - nur schwer
nachvollziehbar, dass die beiden Arten Kyklioacalles
navieresi und Kyklioacalles roboris
von den rheinischen Lokalfaunisten bisher nicht getrennt wurden. Denn
tatsächlich kommen sie im Rheinland sehr häufig „nebeneinander“ vor, auch wenn
sie, wie noch zu zeigen sein wird, verschiedene Habitatansprüche stellen.
Tatsächlich sind die Arten selbst ektoskelettal
leicht zu unterscheiden, wenn man alle
Merkmale einbezieht und davon ausgehen darf, dass an einem Fundpunkt immer nur
eine Art vorkommt. Letzte Sicherheit kann natürlich wie bei allen westpaläarktischen
Cryptorhynchinae (außer den Arten des Genera Echinodera und Ruteria)
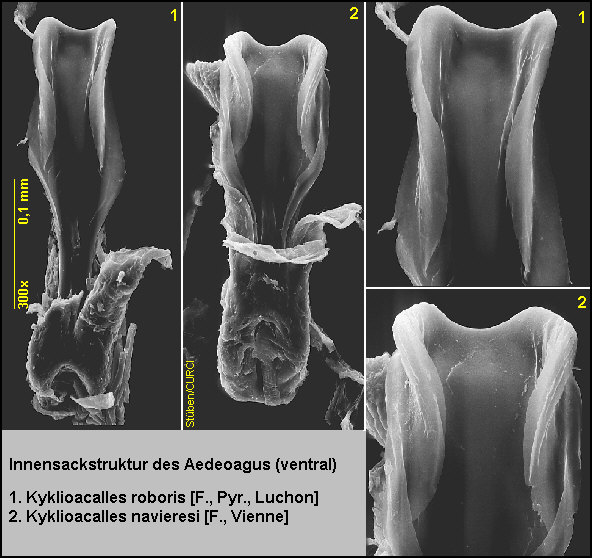
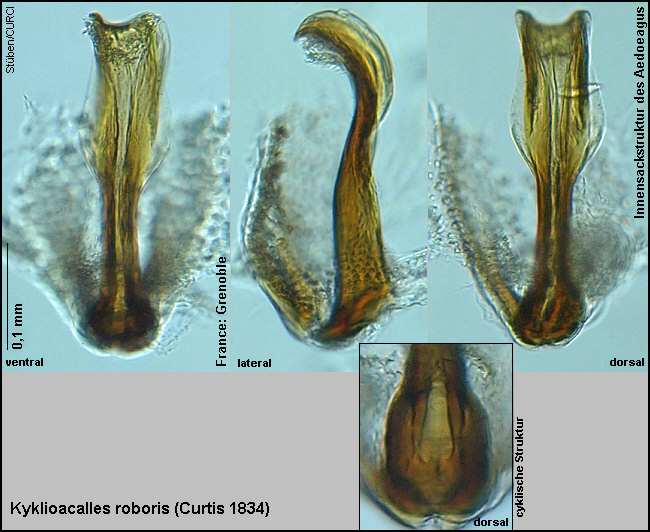
nur die Innensackstruktur des Aedoeagus geben.
Kyklioacalles
navieresi
unterscheidet sich von Kyklioacalles
roboris:
· 1. durch
kürzere Elytren: 1,04x - 1,10x länger als breit; "kastenförmig",
untersetzt, vor der Mitte mit annähernd parallelen Seiten, zur Spitze hin
halbkreisartig verrundet [Fig. W25.3][Fig. W25.4][Fig. W25.5];
· 2. durch
Elytrenstreifen, deren Punktur auch auf den äußersten Streifen kaum größer wird
und hier auf die immer noch deutlich breiteren Intervalle nicht übergreift; Intervalle deutlich flacher [Fig. W25.6];
· 3. durch
das weitgehende Fehlen von Borstenkämmen auf den Intervallen (fehlen immer auf
dem Elytrenabsturz); Intervalle in der Regel nur mit wenigen winzigen,
aufrechten Borsten besetzt [Fig. W25.5];
· 4. durch
das sehr breite, annäherend parallelseitige Pronotum: 1,25x - 1,33x breiter als
lang; breiteste Stelle am Ende des 1. basalen Drittels (auch in der Mitte);
Seiten ± parallel
oder von der breitesten Stelle bis zur Pronotumbasis gradlinig enger werdend [Fig. W25.4];
· 5. durch
die Form des Aedoeagus: Spitze verrundet, ebenfalls Medianlobus seitlich
gleichmäßig langoval verrundet [Fig. W25.7];
· 6. durch
die Innensackstruktur des Aedoeagus: Diese ist 3x länger als (an der breitesten Stelle) breit [Fig. W25.8][Fig. W25.9][Fig. W25.10].
In diesen Merkmalen unterscheidet sich Kyklioacalles roboris von Kyklioacalles navieresi:
· 1*. durch längere Elytren; 1,10x - 1,18x länger
als breit; seitlich langoval verrundet, zur Spitze hin mit einer schwachen
seitlichen Depression (also hier nicht
gleichmäßig, halbkreisartig verrundet) [Fig. W25.11][Fig. W25.4][Fig. W25.5];
· 2*. durch Elytrenstreifen, deren Punktur zu den
äußersten Streifen hin deutlich größer wird, auf die Intervalle übergreift und
diese auf schmale, stark hervortretende Grate reduziert; Intervalle stärker
kielartig hervorgehoben [Fig. W25.6];
· 3*. durch die auf dem 2. und 4. Intervall des
Elytrenabsturzes gut zu erkennenden feinen Borstenkämme [Fig. W25.5];
· 4*. durch ein schmaleres Pronotum: 1,16x - 1,23x
breiter als lang; breiteste Stelle am Ende des 1. basalen Viertels; bis
unmittelbar vor dem Vorderrand seitlich gleichmäßig schwach verrundet. [Fig. W25.4];
· 5*.
durch die Form des Aedoeagus: Spitze abgeflacht; Medianlobus seitlich
vor der Spitze "plötzlich" enger werdend [Fig. W25.12];
· 6*.
durch die Innensackstruktur des Aedoeagus: Diese ist 4x länger als (an der breitesten
Stelle) breit [Fig. W25.13][Fig. W25.9][Fig. W25.10].
Kyklioacalles
roboris wird
ebenso oft aus der Ästchenstreu von Quercus
robur wie auch aus der Streu verschiedener anderer Laubbäume, z.B. Castanea sativa (Fagaceae), gesiebt.
Selbst im Detritus von Carpinus betulus findet
sich diese Art oder kann an schwülen Mai-Tagen sogar von Corylus avellana (Corylaceae) geklopft werden. An Birkengewächsen
(Betulaceae) wird die Art seltener angetroffen, aber der Autor konnte die Art
in den zentralen Pyrenäen an dünnen, im Absterben begriffenen Zweigen und
Stockausschlägen von Fraxinus
(Oleaceae) beim (ektophytischen) Rindenloch- oder Raspelfraß sowie des Nachts
bei der Eiablage beobachten [Fig. W25.14]. Selbst von den Zweigen junger Weißtannen (Abies alba) wurde die Art in größerer Höhe (Austria: Kärnten)
geklopft; und so ließe sich denn diese silvicol-phytodetriticole (manchmal auch
xylophage), aber stets polyphage Bindung anhand einer mehr oder weniger
unspezifischen Reihe von Baum- und Straucharten fast beliebig fortführen. [Fig. W25.15] Dass sich diese
Arten jedoch in verpilzten Zweigen entwickeln, wie manchmal in faunistischen
Arbeiten zu lesen ist, halte ich für ein - wenn sich auch hartnäckig haltendes
- Märchen. [Koch 1992: 301]
Auch bei Kyklioacalles
navieresi scheint mir keine Präferierung von spezifischen Baum- oder
Straucharten vorzuliegen; jedenfalls lassen sich aus den bisherigen
Aufsammlungen (überwiegend Siebungen) auf die habitatspezifischen Bedingungen
bzw. Wirtspflanzenbindungen keine unmittelbaren Rückschlüsse ziehen. Auch diese
Art scheint ausgesprochen oligo- bzw. polyphag zu sein! [Nota]
Es gehört zu den Binsenweisheiten entomologischer Forschung, dass
man auf der Suche nach den habitatsspezifischen Voraussetzungen verschiedener,
nahe verwandter Arten immer das Gesamtverbreitungsgebiet
einer Art – zumal wenn es praktisch die ganze Westpaläarktis umfasst – im
Auge behält. Abgesehen von endemischen Arten oder solchen mit einem sehr
begrenzten Verbreitungsgebiet können lokalfaunistische Arbeiten zu einer
solchen Fragestellung bekanntlich kaum einen nennenswerten Beitrag leisten. Das
unterscheidet das hierzulande immer noch weit verbreitete und häufig
ausschließlich betriebene "Sammeln" in engen, oft sogar politischen
(nicht selten sogar „historisch“ längst überholten) Grenzen von der biogeographischen Forschung [Fig. W25.16].
Insgesamt wurden von mir im Jahre 2002 1095 Exemplare beider Arten
von über 150 Fundorten in ganz Europa untersucht. [Stüben 2003a: 146/148] In
keiner einzigen Gesiebe-Probe fanden sich beide Arten gleichzeitig. Das ist
ungewöhnlich, sollte man doch annehmen, dass bei Siebungen an vielen Plätzen in
einem Radius von mehreren Hundert Metern Aufsammlungen stattfinden. Aber
tatsächlich fanden sich auch in eigenen Aufsammlungen, die häufig sehr
"diversifizierend" (in verschiedenen Habitaten und an
unterschiedlichen Baumarten eines Fundplatzes) angelegt sind, niemals beide
Arten in einer einzigen Probe. Ich gehe daher hier davon aus, dass es sich bei
dem Vorkommen der beiden Arten Kyklioacalles
roboris und Kyklioacalles navieresi nicht
um ein allopatrisches, wohl aber parapatrisches Verbreitungsmuster handelt (die
jeweiligen Populationen also oft im selben geographischen Raum
"mosaikartig" vorkommen, sich aber hier nicht
"überlappen").
Konnten - wie oben beschrieben - bei den beiden
genannten Arten bisher keine Unterschiede in den Bindungen an spezifische Fraß-
und Entwicklungspflanzen festgestellt werden, so scheint dies bei den
abiotischen Anspruchsfaktoren anders zu sein. [Fig. W25.1][Fig. W25.2]
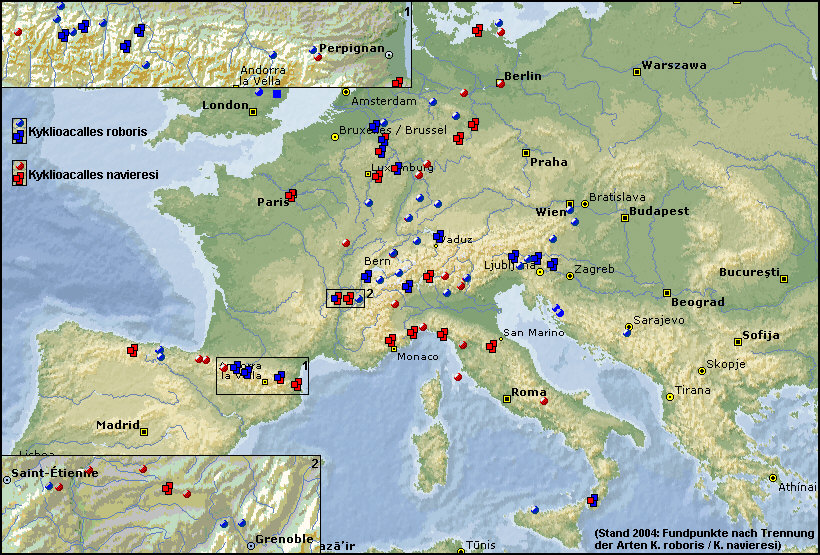
Die Karte [Fig. W25.17] gibt den Stand nach der Artaufspaltung des Kyklioacalles roboris-Komplexes anhand von 1611 Exemplaren und mittlerweile 232 teils durch Koordinaten gestützten Fundorteinträgen bis zum
Ende des Jahres 2004 wieder.
[Fundorte: K. navieresi]
[Fundorte: K. roboris]
In einer ersten Annäherung scheint Kyklioacalles navieresi im Süden Europas (z.B. Pyrenäen bzw. Alpen)
nicht bis in den extremen montanen Raum vorzustoßen. Umgekehrt würde ich bei Kyklioacalles roboris zunächst
vorsichtig von einer auch montanen
Art sprechen, die kälteresistenter zu sein scheint, also durchaus die höheren
Lagen der Alpen und der Pyrenäen erreicht. Zieht man z.B. eine Linie von
Saint-Étienne nach Grenoble, einen Raum, den ich sehr intensiv untersucht habe,
dann stellt man fest, dass auf dem Mont Pilat und in den höheren Lagen um
Grenoble nur die Art Kyklioacalles
roboris gesiebt werden kann, während Kyklioacalles
navieresi eher die mittleren, Rhône-nahen, eindeutig xerothermeren Lagen
bis 300 m präferiert. [Fig. W25.17] Zunächst würde ich also eine temperaturabhängige
(feuchtigkeitsabhängige?) Verbreitung (im Jahresmittel) der beiden Arten als
erste Arbeitshypothese annehmen.
Nähert man sich mit einer solchen
Arbeitshypothese dem Rheinland, ist man beim Betrachten der entsprechenden
Verbreitungskarte zunächst mehr als überrascht und beginnt zu begreifen, warum
Lokalfaunisten sich so schwer getan haben, die Unterschiede zwischen den beiden
Arten wahrzunehmen. Warum hätten sie auch danach suchen sollen? Die Gemengelage
beider Arten scheint keine klaren Präferenzen erkennen zu lassen, das
"Knäuel" sich nicht aufzulösen: [Fig: W25.18]
Es sollte - scheinbar - aber noch "uneindeutiger"
kommen. Bei einer der vielen gemeinsamen Exkursion mit den Kollegen von COLEO
auf dem Bausenberg hatte ich am 21.6.2003 zum ersten Mal beide Arten im Sieb.
Nirgendwo im gesamten europäischen Verbreitungsgebiet war es mir gelungen,
beide Arten an einem ausgewiesen kleinräumigen Standort - der Vulkankegel des
Bausenbergs in der Eifel umfasst gerade einmal
1 km2 - nachzuweisen. [Fig. W25.19] Sollte also dieser kleine Eifelvulkan, der schon 30 Jahre
zuvor eine ganze Generation rheinischer Entomologen beschäftigt hat, mit Recht
als „Schatzkammer der rheinischen Fauna“ bezeichnet wurde und schließlich am
14.4.1981 endgültig unter Naturschutz gestellt wurde [Becker 1975] [Thiele
& Becker 1975][Hoffmann & Thiele 1982], ein weiteres „Geheimnis“
preisgeben?
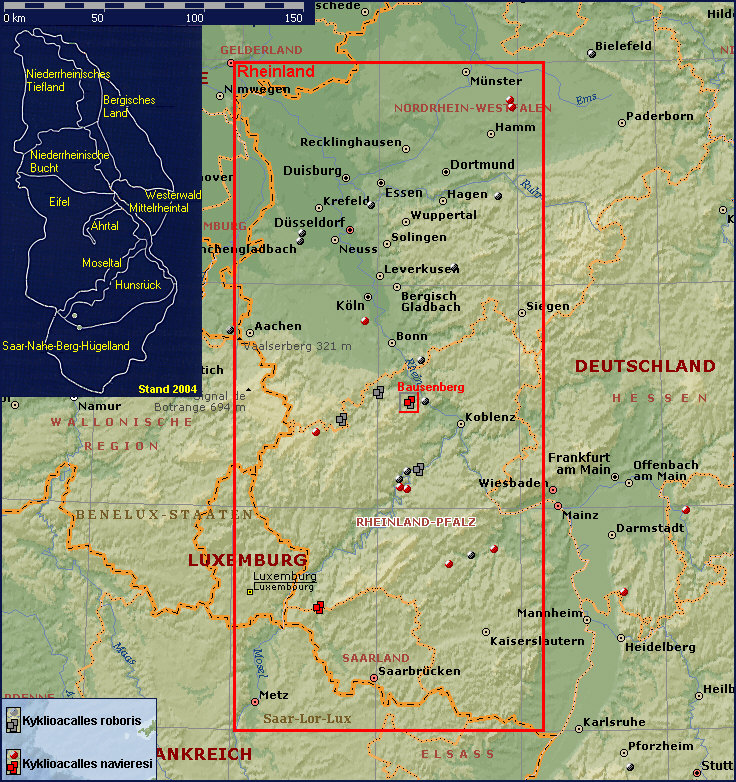
Faunistisch-ökologische Untersuchungen auf dem Bausenberg
Der Bausenberg, der mit einer Höhe von 339,8 m
über N.N. im Regenschatten der Hocheifel liegt (N50°28’ E7°13’), ist vom milden
Klima der Rheineifel geprägt. [Fig. W25.19] Sowohl der zum Brohltal hin abfallende, teils natürliche
belassene, teils vom Abbau der Schlacke bestimmte Südhang als auch der obere
Teil des Ringwalles werden von Schlacken mit geringer Bodenbildung eingenommen.
Dieser Teil kann rasch austrocknen und sich unter der offenen Vegetation, die
jedoch in den letzten 30 Jahren in großen Bereichen verbuschte, stark erwärmen.
Der östliche und westliche Teil des Kraterfußes bestehen hingegen überwiegend
aus schluffigen, meist kalkreichen Böden mit einer äolischen Komponente:
Windeinwehungen aus dem lössreichen Ackerland der Umgebung. Wenigstens hier kam
es zu einer - wenn auch durch die Hangneigung und die nachrutschenden Schlacken
und Aschen nicht ungestörten - Bodenentwicklung. [Stephan 1975]
Die ersten koleopterologisch-faunistischen
Bemerkungen zum Bausenberg finden sich bei C. Roettgen [Roettgen 1911]. Aber
erst die Arbeiten von K. Koch und H. Gräf in den 70er und 80er Jahren des
letzten Jahrhunderts richteten das Augenmerk der rheinischen Koleopterologen
auf den Bausenberg. [Koch 1975] [Koch & Gräf 1982].
Es war die Zeit (1970-1973), in der der Autor noch als Schüler mit
der Erforschung der rheinischen Curculionoidea begann und an den Exkursionen
der Arbeitsgemeinschaft Rheinischer Coleopterologen unter der Leitung seines
späteren Lehrers A.M.J. Evers (Krefeld) auf dem Bausenberg teilnahm. Die
damaligen Exkursionen standen ganz im Zeichen des Projekts zur „Erforschung der
Tier- und Pflanzenwelt des Bausenbergs“, wie es von H. U. Thiele und J. Becker
initiiert wurde. [Thiele & Becker 1975] [Becker 1975] [Hoffmann &
Thiele 1982]
Fundumstände von Kykliocalles navieresi und
Kyklioacalles roboris am Bausenberg [Fig. W25.20]
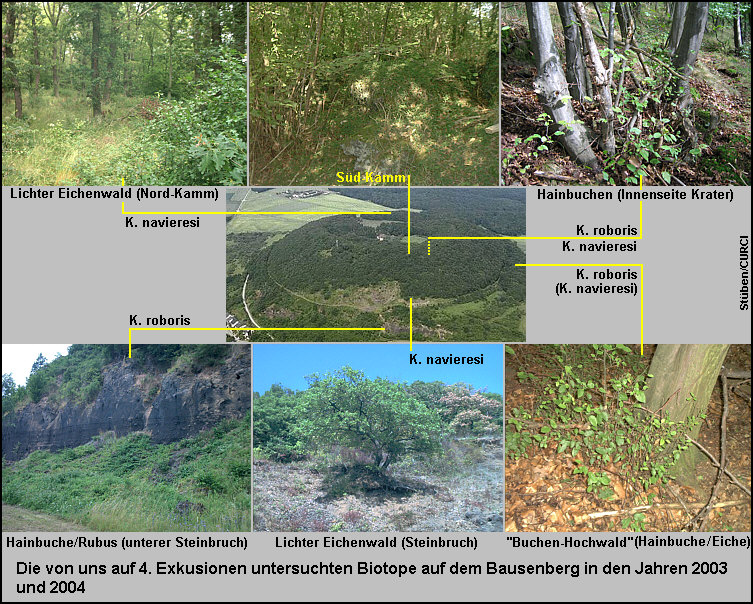
Nachdem ich am 21.6.2003 während eines kurzen
Aufenthalts mit der „Gemeinschaft für Coleopterologie“ (COLEO e.V.) am
Bausenberg beide Arten siebte, fand zusammen mit Friedhelm Bahr (Viersen) am
21.9.2003 eine zweite Begehung der Süd-Seite des Kraterkamms und der nach
Norden weisenden Innenseite des Kraters statt. Dabei wurde von mir an solitären
Eichen sehr trockener Standorte in der Nähe des Steinbruchs Kykliocalles
navieresi [Fig. W25.21] und von Friedhelm Bahr an Hainbuchen auf der Innenseite des
Kraters, einem etwas feuchteren Standort [Fig. W25.22], Kyklioacalles roboris in jeweils drei Exemplaren
gesiebt. Siebungen an Haselnusssträuchern ergaben keine weiteren Nachweise.
Am 10. und 11.7.2004 besuchten wir dann ein
drittes Mal die Trockenhänge der Süd-Seite und diesmal die östlich bzw.
nördlich gelegenen Hochwälder am Rande des Kraters. Es hatte in den Wochen und
Tagen zuvor heftig geregnet, und die Temperatur betrug tagsüber 15 – 17 °C. Aus
einem Eichenwald auf dem Nordkamm siebten wir in großer Anzahl ausschließlich Kykliocalles
navieresi. [Fig. 25.23] Ebenso fand ich diese Art erneut an solitären Eichen sehr
trockener Standorte in der Nähe des Steinbruchs auf den Hängen der Südseite in
über 280 m über N.N. [Fig. 25.24] Diese zum Brohltal abfallenden Südhänge sind thermisch sehr
begünstigt und weisen zahlreiche xerotherme Gehölze auf hohlraumreichen
Schlackeböden auf. [Stephan 1975] An dem zuletzt genannten Fundort fand
ebenfalls eine Nachtexkursion bei ca. 13 °C statt. Trotz intensiven Klopfens
konnten nur wenige Acalles dubius-Exemplare - eine auf dem Bausenberg
sehr häufige Art - vor allem an den Stockausschlägen von Hainbuchen (Carpinus) nachgewiesen werden.
Völlig anders präsentierte sich diesmal jedoch die
Artenzusammensetzung an den Hainbuchen-Standorten. Wieder fand sich wie schon
im Jahr zuvor auf dem etwas feuchteren Hainbuchen-Standort der nach Norden
weisenden Innenseite des Kraters Kyklioacalles roboris zusammen mit
zahlreichen Exemplaren der feuchte Standorte präferierenden Art Ruteria
hypocrita. Doch im gleichen Gesiebe fanden sich auch Exemplare von Kykliocalles
navieresi! Die gleiche Artenzusammensetzung stellte sich in den Siebungen
am Rande eines feuchten Buchen-Hochwaldes am Fuße des Osthanges ein: Neben der
feuchte Standorte präferienden Art Ruteria hypocrita wurde ein Exemplar
von Kyklioacalles roboris und zwei weitere Exemplare von Kykliocalles
navieresi nachgewiesen. [Fig.W25.25] Den gleichzeitigen Fund beider Kyklioacalles-Arten führe
ich jedoch darauf zurück, dass es an diesen Standorten immer auch solitäre
Eichen gibt. [Fig. W25.26] So konnten infolge äolischer und fluvialer
Stöckchenverdriftungen auf steiler Hanglage zu diesem Zeitpunkt noch keine
streng getrennten Siebungen durchgeführt und damit getrennte Nachweise erzielt
werden. Die Bindung von Kyklioacalles roboris an feuchtere Standorte
präferierende Hainbuchen-Bestände auf dem Bausenberg fand jedoch eine
unmittelbare Bestätigung in den tieferen Lagen des Südhangs unter 250 m über
N.N. Dieser Bereich ist heute weitgehend verbuscht und bei weitem nicht mehr so
xerotherm wie die weitläufigen Eichenbestände oberhalb des Steinbruchs. Breite
Hecken mit vereinzelten Hainbuchen werden von landwirtschaftlich genutzten
(Trocken-)Wiesen unterbrochen. Hier ist es möglich gezielt an solitären
Hainbuchen zu sieben ohne Gefahr zu laufen, auf von Wind und Wetter
verdriftetes Eichensubstrat zu stoßen. [Fig. W25.27] Und an diesen oft von dichten Rubus-Gewächsen eingefassten Hainbuchen auf einem relativ flachen
Terrain fand sich dann tatsächlich nur Kykliocalles roboris.
Solche Indizien reichen natürlich nicht aus um
sicherzustellen, dass die genannten Arten sich nach Wirtspflanzen „aufspalten“
lassen – hier Kykliocalles roboris an Carpinus betulus und Kykliocalles navieresi an Quercus
robur -, und dass dieser zunächst nur für den Bausenberg spezifischen Nischenbesetzung an unterschiedlichen
Gehölzen eine ökologische Priorität eingeräumt werden darf. Warum sollte z.B.
die hygrophilere Art Kykliocalles roboris nicht auch an
feuchteren Quercus-Standorten
vorkommen?
Methodisch ist
solchen Fragenstellungen bei Cryptorhynchinae ganz sicher nicht mit
Fallen-Aufsammlungen [Becker 1975] oder gar mit dem Klopfschirm beizukommen.
Beide Fangmethoden haben sich nach jahrelangen und äußerst intensiven
Aufsammlungen in den 70er Jahren auf dem Bausenberg als wenig ergiebig bzw.
aussagekräftig erwiesen: So nennt Koch lediglich 5 Nachweise für Acalles
dubius (ehemals Acalles turbatus) und zählt gerade einmal 4
Exemplare von Ruteria hypocrita (ehemals Genus: Echinodera) auf. Kyklioacalles
roboris (hier noch beide Arten!) käme danach mehrfach, aber meist
vereinzelt vor, sei also keineswegs „häufig“. [Koch 1975:294]
Dass bei uns in nur
einer einzigen, 15 Minuten währenden Siebung auf gerade einmal einem Hektar ein
Vielfaches der genannten Exemplare sich auf dem Bausenberg einstellte, zeigt
nur, dass Klaus Koch schon damals mit seiner Einschätzung der Relevanz der
eingesetzten Fangmethoden sehr selbstkritisch umzugehen wusste: „Eine
Sammelmethode, die sicherlich noch wesentlich zur Bereicherung der Artenliste
(des Bausenbergs – der Verfasser) beigetragen hätte, nämlich der Gebrauch des
Käfersiebes, wurde leider nicht angewandt.“ [Koch 1975: 280]
Man könnte es an
dieser Stelle auch zeitgemäßer formulieren: Cryptorhynchinae haben hierzulande auf Roten Listen so lange
nichts zu suchen, wie die geeignete Fangmethode, nämlich die „Siebung“, nicht
oder nur spärlich bzw. gelegentlich zur Anwendung kommt!
Darüber hinaus ist die kontinuierliche,
durch keine längeren Brachzeiten infolge von Rodungen unterbrochene Waldgenese selbst der
entscheidende Faktor für den Nachweis bzw. Nicht-Nachweis heimischer Acalles-Arten.
Wo der Wald einige Jahre oder Jahrzehnte großräumig Viehweiden Platz machen
musste, werden sich keine stabilen Populationen mehr nachweisen lassen (auch
nicht in Jahrhunderten!). Anders formuliert: Der Nachweis zahlreicher Acalles-Arten ist ein hervorragender
Indikator für eine lang anhaltende, ununterbrochene Waldgenese.
Umgekehrt sagt das Fehlen flugunfähiger Cryptorhynchinae nichts über für Acalles-Arten
heute durchaus geeignete Waldbiotope aus. Damit kommt der „Seltenheit“ bzw. dem
Noch-Vorhandensein dieser Arten – über deren
Schutz hinaus (das ist aber ein anderes Thema) – natürlich auch keine weiterführende Bedeutung als
Indikator für schützenswürdige Waldstandorte a priori zu. Wo Cryptorhynchinae fehlen, sind naturbelassene
Waldstandorte nicht weniger schützenswert!)
Die am 25.9. 2004 mit dem Kollegen Horst-Dieter Matern
vorgenommenen Siebungen auf der nach Norden weisenden Innenseite des Kraters -
diesmal an gezielt ausgewählten einzelnen Hainbuchen und Eichen - ergab das
bekannte Bild: Obwohl sehr genau darauf geachtet wurde, dass diese letzten
Siebungen unmittelbar im Stammbereich der Hainbuchen stattfanden und von den
weiter oberhalb angepflanzten Eichen über äolische
und fluviale Verdriftungen kein Material eingetragen wurde, fanden sich
an diesem Standort an den Hainbuchen wieder beide Arten! [Fig. W25.28]
Offensichtlich ist der Gradient „trocken-feucht“ auf den
porösen Schlacken dieses Standortes so „grenzwertig“, dass selbst mit
kleinräumigen Siebungen die Arten nicht mehr gezielt gesammelt bzw. getrennt
werden können. Allerdings ist unter 100en von Aufsammlung in ganz Europa dies
nachweislich der einzige Standort,
an dem beide Arten kleinräumig nebeneinander vorkommen. Keineswegs aber stellt
dieser eine Fall eine zu
„vernachlässigende Größe“ dar – im Gegenteil: Er liefert den wichtigen Hinweis,
dass offensichtlich die Bindung an Laubbaumarten die eigentliche, zu „vernachlässigende Größe“ bei diesen und anderen
mitteleuropäischen Waldarten aus der Unterfamilie der Cryptorhynchinae ist.
Die These, dass in weiten Teilen Europas Kyklioacalles
roboris die feuchtigkeitsresistentere Art ist, muss daher um die
These, dass Kyklioacalles
navieresi die eindeutig xerothermere Standorte präferierende Art
ist, ergänzt werden. So fand sich auf den Südhängen des Bausenbergs (am Steinbruch)
in den lichten, trockenen Quercus robur
–Wäldern (besser: -Gebüschformationen) ausschließlich diese Art! Dieser Teil des Bausenbergs kann rasch austrocknen und sich
unter der offenen Vegetation in den Sommermonaten sehr stark erwärmen. Ich wage
daher die Prognose, dass man auf solchen Standorten niemals Kyklioacalles roboris finden wird. Legt
man die Erfahrungen mit anderen Kyklioacalles-Arten
zugrunde, werden sich die Larven dieser Art - während der dreimonatigen
Entwicklung im Hochsommer - in den ersten beiden Larvalstadien mit solchen
trockenen Habitatstrukturen nicht abfinden, hier also keine geeigneten
Entwicklungsmöglichkeiten vorfinden (vgl. dazu: [Stüben 2003e, 2003d, 2004g].
Natürlich sollten derartige im Freiland gewonnene Erfahrungen
und unter den genannten methodischen Bedingungen aufgestellte Thesen immer durch Laboruntersuchungen ergänzt
werden. Nur so lassen sich ökologische Feldstudien auf ihre biologischen Wurzeln hin überprüfen –
vom „Kopf auf die Füße stellen“! Dabei könnten solche Untersuchungen ihren
Ausgang in der Nachzucht der jeweiligen Art nehmen. Während jedoch bei den
Imagines eine hohe Toleranzschwelle anzunehmen ist, sollten die Larven in ihren
jeweiligen Stadien Feucht-Trocken-Experimente durchlaufen bzw. solchen
Experimenten ausgesetzt werden.
Das wäre dann aber weiterführenden Studien vorbehalten.
Literatur
Becker, J. (1975): Art und Ursache der
Habitatbindung von Bodenarthropoden (Carabidae - Coleoptera, Diplopoda,
Isopoda) xerothermer Standorte in der Eifel. - Beitr. Landespflege Rhld.-Pfalz
(Oppenheim), Beiheft 4, 89-140.
Begon, M. & Harper, J.L. & Townsend, C.R.
(1991): Ökologie: Individuen, Populationen und Lebensgemeinschaften, 1024 S.,
Basel / Berlin.
Hoffman, H.-J. & Thiele, H.-U. (Hrsg. 1982): Neue
Untersuchungen zur Tierwelt des Bausenbergs in der Eifel. – Decheniana -
Beihefte, 27: 1 – 279, Bonn.
Koch, K. (1975): Untersuchungen an der
Koleopterenfauna des Bausenbergs (Eifel). - Beitr. Landespflege Rhld.-Pfalz
(Oppenheim), Beiheft 4, 274-325.
Koch, K. & H. Gräf (1982): Nachtrag
zur Koleopterenfauna des Bausenbergs (Eifel). - Decheniana-Beihefte (Bonn) 27,
241- 254.
Koch, K. (1992): Die Käfer Mitteleuropas -
Ökologie. - In: Freude,
H. ; Harde, K. & Lohse, G.H. (Hrsg.): Die Käfer Mitteleuropas, Bd. E3: 389 S., Krefeld.
Roettgen, C. (1911): Die Käfer
der Rheinprovinz. – Verh. Naturh. Ver. preuß. Rheinl. Westf. u. Reg. Bez. Osnabrück, 56: 345 S., Bonn.
Sprick, P. & Stüben, P.E. (2000): Ökologie der kanarischen Cryptorhynchinae
außerhalb des Laurisilva. - Cryptorhynchinae-Studie 11 - In: Stüben, P.E.
(2000b), SNUDEBILLER 1. Die
Cryptorhynchinae der Kanarischen Inseln (CD ROM): 318-341,
Mönchengladbach.
Stephan, S. (1975): Die
Vegetationsverhältnisse am Bausenberg in der Eifel. – In: Thiele, H.-U. &
Becker, J. (Hrsg. 1975): Der Bausenberg - Naturgeschichte eines
Eifelvulkans. - Beitr. Landespflege Rheinland-Pfalz (Oppenheim) Beiheft 4,
57-78.
Stephan, S. (1975): Die Böden
am Bausenberg in der Eifel. – In: Thiele, H.-U. & Becker, J. (Hrsg. 1975):
Der Bausenberg - Naturgeschichte eines Eifelvulkans. - Beitr. Landespflege
Rheinland-Pfalz (Oppenheim) Beiheft 4, 33-56.
Stüben, P.E. (1997): Rüsselkäferzönosen in aufgelassenen
Ton- und Sandgruben. Anmerkungen zu Mosaik-Sukzessionen bei der Nutzung des
geplanten Naturschutzgebietes Holter Heide bei Brüggen (Col. Curculionoidea),
in: Decheniana-Beihefte (Bonn) 36, S. 185-216.
Stüben, P. E. (2003a): Revision des Genus Kyklioacalles
und Beschreibung der Untergattung Palaeoacalles subg. n. unter
Heranziehung phylogenetischer, morphogenetischer und biogeographischer Aspekte
(Curculionidae: Cryptorhynchinae). - SNUDEBILLER 4, Studies on taxonomy,
biology and ecology of Curculionoidea, Mönchengladbach: CURCULIO-Institute,
116-166.
Stüben P.E. (2003d): Breeding of Kyklioacalles
euphorbiophilus Stüben 2003 (Coleoptera: Curculionidae: Cryptorhynchinae)-
Weevil News: http://www.curci.de/Inhalt.html, No. 15: 6 pp.,
CURCULIO-Institute: Mönchengladbach. (ISSN 1615-3472).
Stüben P.E. (2003e): Zucht von Kyklioacalles
euphorbiophilus Stüben 2003 (Coleoptera: Curculionidae: Cryptorhynchinae) -
COLEO: http://coleo.de/2003/Kyklio/KyklioZucht.html, Nr. 4: S.
7-16, (ISSN 1616-3281).
Stüben, P.
E. (2004g): Zur Biologie von Acalles poneli Stüben 2000 -
(Coleoptera: Curculionoidea). - Weevil News: http://www.curci.de/Inhalt.html,
No. 19: 13 pp., CURCULIO-Institute: Mönchengladbach (ISSN 1615-3472).
Stüben, P. E. (2005e, in print):
Beschreibung neuer Cryptorhynchinae aus dem mediterranen Raum (Coleoptera:
Curculionidae: Cryptorhynchinae). - SNUDEBILLER 6, Studies on taxonomy, biology and ecology of Curculionoidea,
Mönchengladbach: CURCULIO-Institute.
Thiele, H.-U. &
Becker, J. (Hrsg. 1975): Der Bausenberg - Naturgeschichte eines Eifelvulkans.
- Beitr. Landespflege Rheinland-Pfalz (Oppenheim) Beiheft 4, 1-394.
*
Die Redaktion der WEEVIL NEWS des CURCULIO-Instituts bedankt sich bei COLEO e.V.
für die Rechte am Wiederabdruck des Artikels und – für diesen Zweck – für
die Rechte an den Abbildungen.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}