Weevil News
|
No. 31 |
13 pp. |
2. October 2005 |
ISSN 1615-3472
|
|
|
Stüben, P. E. (2005): “Basar
Taxonomie”? – Ein erfolgreiches Kreuzungsexperiment zu Acalles aeonii
Wollaston, 1864 (Coleoptera: Curculionidae: Cryptorhynchinae). - Weevil News: http://www.curci.de/Inhalt.html,
No. 31: 13 pp., CURCULIO-Institute: Mönchengladbach. (ISSN 1615-3472). |
|||||
"Basar Taxonomie"?
Ein
erfolgreiches Kreuzungsexperiment zu Acalles aeonii Wollaston, 1864* (Coleoptera: Curculionidae:
Cryptorhynchinae)
von
P.E. Stüben, Mönchengladbach
Mit 31
Abbildungen
Abstract
As a result of a successful cross-breeding of Acalles aeonii
Wollaston 1864 x Acalles bodegensis Stüben 2000 the following
synonymy is established definitively: Acalles aeonii Wollaston 1864 =
Acalles bodegensis Stüben 2000 syn. nov. A detailed report on
rearing and cross-breeding of the former two species on its host plant Aeonium holochrysum W. & B. during a number of
generations is given and the results are discussed. With a detailed
discussion to the necessity of cross-breeding experiments in the field of
taxonomic-systematic research. Including 31 colour photographies.
Key words
Coleoptera, Curculionidae, Cryptorhynchinae, Acalles aeonii, Acalles
bodegensis, taxonomy, biology, ecology, host plant, breeding, synonym,
Canary Islands, Tenerife.
Zusammenfassung
Als Ergebnis
eines aufwendigen, aber erfolgreichen Kreuzungsexperimentes von Acalles
aeonii Wollaston 1864 x Acalles bodegensis Stüben 2000 wird folgende Synonymie endgültig
bewiesen: Acalles aeonii Wollaston 1864 = Acalles bodegensis Stüben
2000 syn. nov. Zucht und
Kreuzung der genannten Arten werden über zahlreiche Generationen hinweg an der
Entwicklungspflanze Aeonium holochrysum W. & B. detailliert beschrieben und die
Ergebnisse diskutiert. Mit einer ausführlichen Diskussion zur Notwendigkeit von
Kreuzungsexperimenten in der taxonomisch-systematischen Forschung.
1. Einleitung
Erster Exkurs
Taxonomen sind bekanntlich keine Systematiker –
jedenfalls in den seltensten Fällen. Ihnen reichen die Regeln der Nomenklatur
in der jeweils neuesten Fassung [ICZN 2000], eine genaue Beobachtungsgabe und
ein nicht näher zu spezifizierendes „Gefühl für Evidenzen und Signifikanzen“.
Während die einen nach dem phylogenetischen System suchen, beschreiben
die anderen gelegentlich Arten und Gattungen was das Zeug hält, ohne nach den
evolutionären Zusammenhängen zu fragen.
Taxonomen sind aber auch keine Biologen – jedenfalls in den seltensten Fällen. In musealer Eintracht mit ihren Präparaten suchen sie nach Merkmalen, schreiben kleine Differentialdiagnosen und bemühen sich irgendwie um eine korrekte Klassifikation ihrer Präparate – nur ganz selten lassen sie das „Leben“ selbst entscheiden.
Ist es nicht erschreckend, sich gelegentlich dabei zu beobachten, wie mit Kollegen um die Signifikanz von Merkmalen und Merkmalsausprägungen geradezu gefeilscht wird, dem "Basar Taxonomie" ein höherer Stellenwert (besser: mehr Aufmerksamkeit) eingeräumt wird als der phylogenetischen Systematik oder gar der biologischen Forschung? Was ist einem die Neubeschreibung eines Kollegen eigentlich wert, dem man in einer Anmerkung, in zwei oder drei Sätzen oder einfach nach einem "Gleichheitszeichen (=)" mal schnell seine Arten (weg)synonymisiert?
Immer wieder geht es dabei um Reglementierungen, um Regeln zur formalen Klassifikation von Taxa, um Namen, Prioritäten und Stabilitäten – oft aber auch um Luftschlösser und Eitelkeiten. Oder darf man die Bemerkung eines geschätzten Kollegen anders verstehen, der meiner besorgten Bitte, er möge mir bei der Abtrennung einer Art behilflich sein, entgegnete: „Die Regeln implizieren, dass mit der Einführung eines neuen Namens für ein Taxon mindestens ein neues Merkmal benannt werden muss. Das aber kann ja wohl nicht so schwer sein - Apomorphien und Plesiomorphien hin oder her!“
Nicht er, sondern ich trage die Verantwortung für diesen Blödsinn: Die Art tatsächlich beschrieben zu haben. Oder ist es nicht unbefriedigend, jahrelang das unbestimmte Gefühl zu haben, man habe in der Vergangenheit eine Art beschrieben, die keinen Platz in der Zufallsgeschichte der Evolution hat?
Damit wäre ich beim Thema, wenn es da nicht noch ein zweites – eher erkenntnistheoretisches - Missverständnis auszuräumen gilt.
Zweiter Exkurs
Weder die morphologische Ähnlichkeit noch die
genetische Distanz sind universell einsetzbare Kriterien, um Arten zu
unterscheiden. Die Reversibilität bzw. Irreversibilität der genetischen Distanz
zwischen ähnlichen Organismen kann und sollte immer noch daran festgemacht
werden, ob sich natürliche Populationen miteinander kreuzen oder nicht. Das ist
der biologische Artbegriff von Mayr [Mayr 1942, 1969], der selbst dort nicht
hinfällig wird, wo auf die Schwierigkeiten von Kreuzungsversuchen und
auf die in der Praxis angeblich nur schwer zu realisierenden experimentellen
Überprüfungen hingewiesen wird. Der einzige Beweis für die Zugehörigkeit zu
derselben Art bei bisexuellen Organismen besteht in der Beobachtung der Fortpflanzung
selbst, und nur ein "Kreuzungsexperiment kann das Ausmaß der reproduktiven
Isolation (endgültig - d. Verf.) klären" [Wägele 2001].
Alles andere sind Regelwerke, Indizien und Ad-hoc-Hypothesen, aber keine "Beweise". Solche Hypothesen sind in einem hohen Maße anfällig, beruhen z.B. auf den Annahmen, dass "subjektiv" wahrgenommene Morphologieunterschiede existieren, Zwischenformen bisher nicht nachgewiesen wurden, spezifische Kopulationsapparate (Schlüssel-Schloss-Prinzip) vorhanden sind oder ein 16S-rRNA-Sequenzunterschied von mindestens 1,5 bis 2 Prozent ausreicht, um eine neue Art zu „begründen“.
Alle diese Regelwerke, Indizien und Hypothesen beruhen auf Konventionen, von denen wir in den seltensten Fällen überhaupt wissen, ob sie sich in der Praxis bewährt haben! Ich schätze, dass auf 500 - 1000 Neubeschreibungen bei den Curculionoidea vielleicht gerade einmal ein Kreuzungsexperiment zur Abstützung differentialdiagnostischer Bemühungen herangezogen wird, um aus der Art-"Entdeckung" eine "Art“ zu machen (wobei ich mir bewusst bin, dass eine Art keine Entität, sondern stets ein "Vorgang" ist). Hingegen ist mir nicht eine einzige Synonymisierung bekannt, die auf einem Kreuzungsexperiment beruht; und dies, obwohl kaum ein Kollege und kein einziger Fach- und Lehrbuchautor darauf verzichten würde, genau ein solches Experiment als die ’ultima ratio’ anzupreisen (ohne nicht umgehend und oft noch im selben Gedankengang darauf hinzuweisen, dass dies in der entomologischen Praxis "ja wohl kaum zu realisieren" sei). Zwischen Theorie und Praxis, zwischen Anspruch und Wirklichkeit klafft hier ein riesiger Graben.
Damit bin ich nun
endlich beim Thema: Denn Acalles bodegensis Stüben 2000 von Tenerife
schien mir gleich nach der Erstbeschreibung eine solche regelkonforme
Indizien-Art zu sein – ein Luftschloss? [Stüben 2000k: 38-39]
Aber
zunächst überwog die Zuversicht, dass die genannten Merkmale, aber vor allem
die Merkmalsunterschiede zu Acalles aeonii Wollaston 1864 in der
Differenzialdiagnose ausreichen würden, hier von einer eigenständigen Art zu
sprechen. Erst 2003 kamen mir bei der Aufstellung des „2. Analytischen Katalogs
zu den Cryptorhynchinae der Westpaläarktis“ erhebliche Zweifel:
"Heute bin ich mir nach nochmaliger Sichtung des gesamten
Materials nicht mehr so sicher, ob es sich bei Acalles bodegensis tatsächlich
um eine eigenständige Art handelt. Die Art müsste am Locus typicus (Tenerife:
Lomo Las Bodegas) noch einmal in Anzahl nachgesammelt werden und sowohl die
Larven, Puppen als auch Imagines mit der Nachbarart Acalles aeonii -
möglichst unter Heranziehung moderner Methoden der DNA-Sequenzierung -
verglichen werden." [Stüben & Behne & Bahr 2003: 23]
Aber will man nicht wieder in eine weitere "Konventionen-Falle" laufen, sollte man molekularbiologische Überlegungen zur Artdifferenzierung zunächst beiseite schieben, denn diese können immer nur eine notwendige, niemals jedoch hinreichende Voraussetzung für Artdifferenzierungen sein. Es gibt keinen absoluten genetischen Distanzwert und kein Kriterium, das den intraartlichen Variationsraum (über viele Populationen hinweg) von dem interartlichen Differenzierungsraum a priori - und nur darum geht es! - unterscheiden könnte (dazu [Stüben 2004e: 101]). Mit anderen Worten: "Ein absoluter (genetischer) Distanzwert kann als Kriterium für die Existenz einer Fortpflanzungsbarriere nicht angegeben werden." [Wägele 2001]
Was bleibt, ist das Kreuzungsexperiment und die einzigartige wissenschaftliche Ungewissheit, wie es ausgehen könnte.
2. Kreuzzucht:
Acalles aeonii x Acalles
bodegensis
Beschreibung der
Ausgangssituation
Aus der Erstbeschreibung
von Acalles bodegensis: „Die neue Art (Acalles bodegensis - d. V.) wurde in Anzahl im Sukkulentenbusch bei Lomo de Las
Bodegas (östliches Anaga) in 500 m Höhe vor allem des Nachts ausschließlich von
Aeonium holochrysum W. & B., einem Halbstrauch mit verzweigten, 1-3
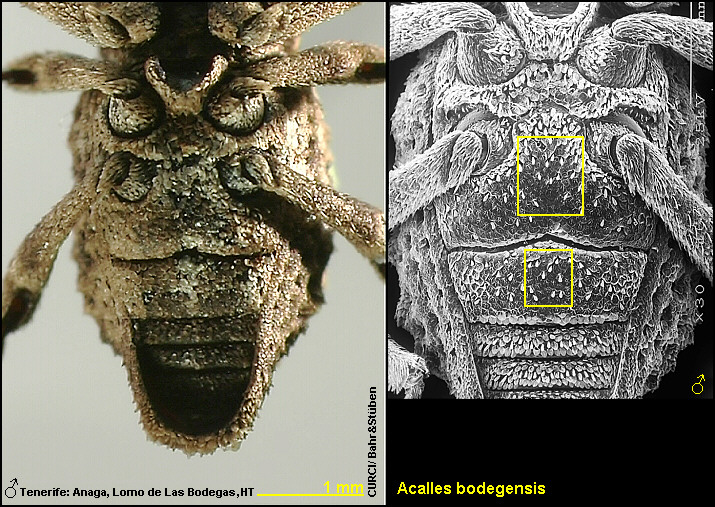
cm dicken, glatten Ästen, geklopft [Fig. W31.1]. Acalles bodegensis scheint nur im äußersten
Nordosten des Anaga-Gebirges vorzukommen. Weiter westlich, sicher aber zwischen
Taganana und Batán de Abajo (oder auf den Südhängen, westlich der Linie El
Bailadero - San Andrés) kommt die Art nicht vor und wird hier von Acalles
aeonii abgelöst. Ein sympatrisches Auftreten
dieser beiden Arten konnte nicht beobachtet werden.“ [Stüben 2000k: 38]
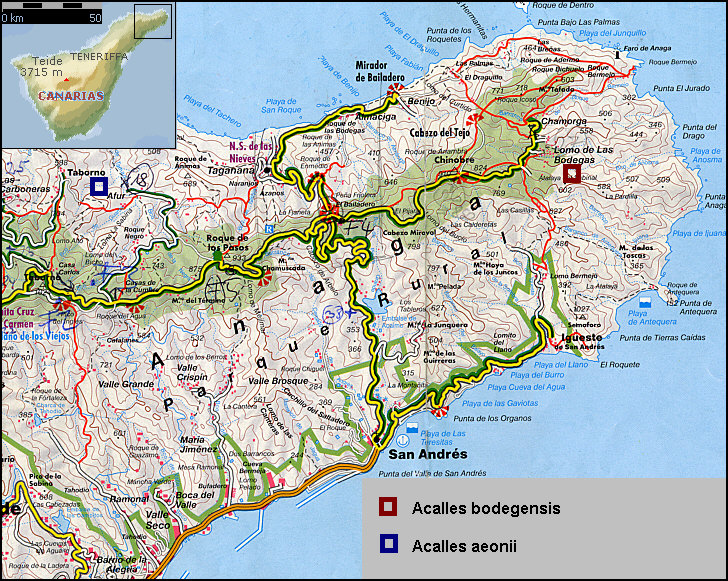
Die genaue Beschreibung der unterschiedlichen Areale und Pflanzenbindungen in der (Re)Deskription von Acalles bodegensis und Acalles aeonii [Stüben 2000e] wurden von Christoph Germann (CH-Bern) und mir Ende Dezember 2003 zum Anlass genommen, umfangreiche Aufsammlungen beider Arten für eine geplante Kreuzzucht in den Insektarien des CURCULIO-Institutes vorzunehmen. Die beiden Fundorte liegen 10 km Luftlinie auseinander und sind durch zahlreiche tiefe Täler und sehr hohe Gebirgskämme getrennt [Fig. W31.2]:
Fundort (Locus typicus) von Acalles bodegensis an Aeonium holochrysum W.
& B.:
27 Ex.: "E., East Anaga
Mts, 19 km NE La Laguna, Bodegas, N28°33'43"W16°09'25", 500m,
22.12.2003, Sukkulentenbusch: Aeonium holochrysum W. & B., leg. Stüben (FO 9)". [Fig. W31.3]
Fundort von Acalles aeonii an Aeonium cuneatum W. & B.:
35 Ex: "E., Tenerife,
Anaga Mts., 10 km NE La Laguna, Afur,N28°33'10"W16°14'59", 300m,
29.12.2003, Sukkulentenbusch: Aeonium cuneatum W.
& B., leg. Stüben (FO 26)". [Fig. W31.4]
Alle Exemplare beider Arten (dazu wurden einige Tiere abgetötet) wurden nochmals im CURCULIO-Institut (Mönchengladbach) mit der jeweiligen Deskription, insbesondere mit der Differenzialdiagnose in der Erstbeschreibung von Acalles bodegensis verglichen bzw. (nach)determiniert. Wieder fanden sich keine morphologischen Abweichungen bei der Zuordnung der Exemplare - insbesondere nach der Form der Innensackstruktur des Aedoeagus - zur jeweiligen Deskription bzw. Redeskription. "Zwischen-" oder "Übergangsformen" konnten nicht festgestellt werden. Die Tiere waren - leicht und schnell - eindeutig einer der beiden Arten zuzuordnen!
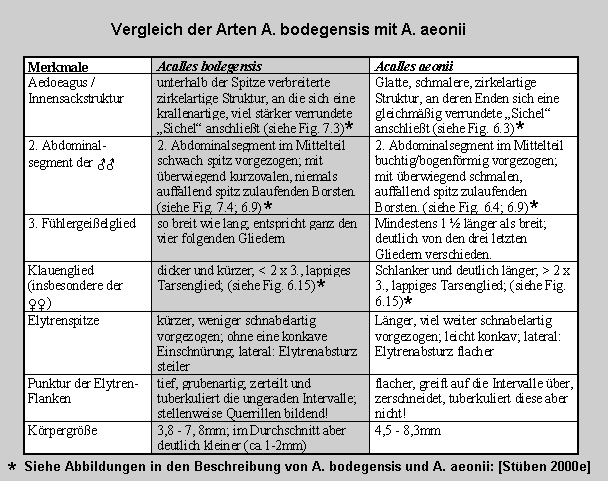
Aus der Differenzialdiagnose in der Erstbeschreibung von Acalles bodegensis:
"Vergleich der Arten Acalles bodegensis und Acalles
aeonii: [Fig.
W31.5]
* Abbildungen zur Tabelle:
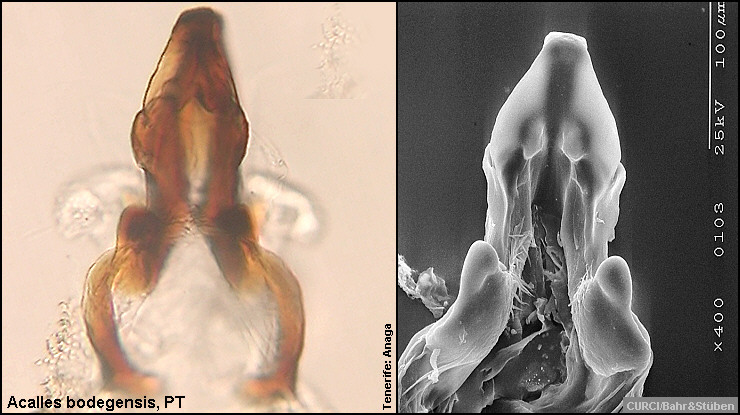
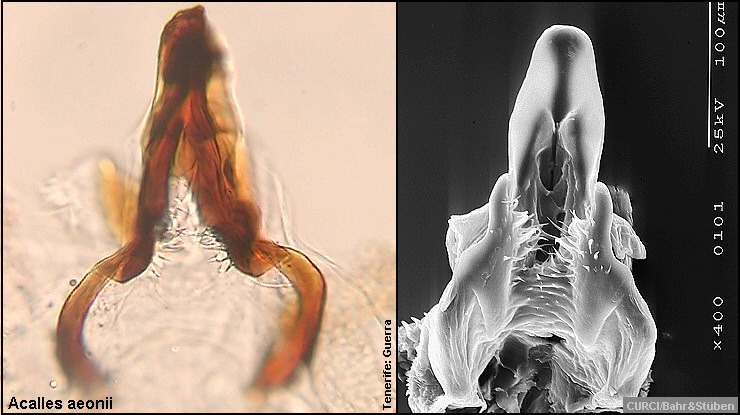
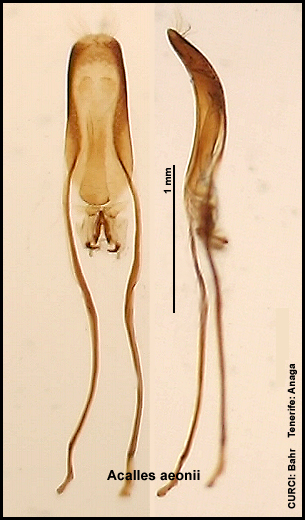
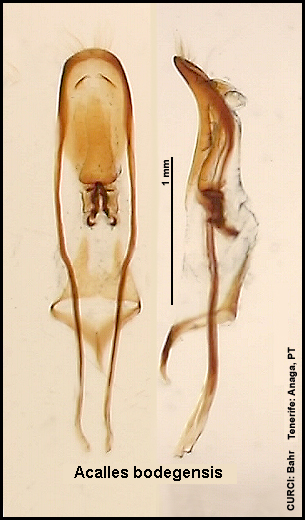
Innensackstruktur des Aedoeagus:
Acalles bodegensis [Fig. W31.6]
Acalles aeonii [Fig.
W31.7]
2. Abdominalsegment:
Acalles bodegensis [Fig. W31.8]
[Fig. W31.9]
Acalles aeonii [Fig.
W31.10] [Fig. W31.9]
Länge des Klauengliedes: [Fig. W31.11]"
Im
deutsch-spanischen Schlüssel aus dem Jahre 2000 heißt es außerdem [Stüben
2000k: 93]:
"20 2.
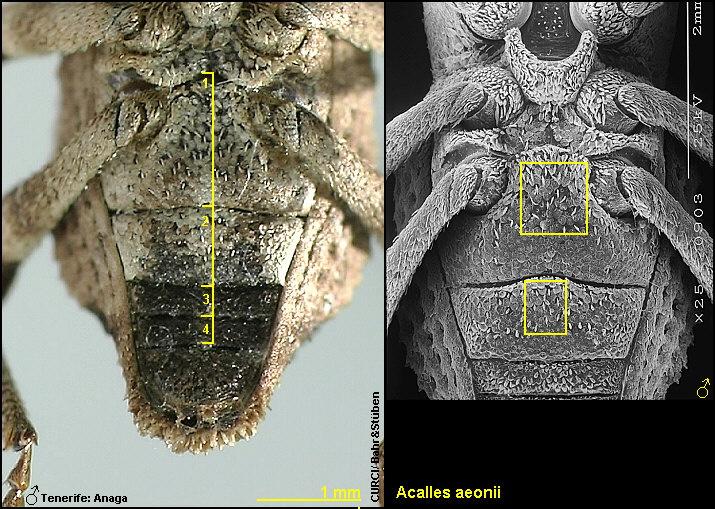
Abdominalsegment mit überwiegend schmalen, auffallend spitz zulaufenden Borsten
[Fig. W31.10]; Klauenglieder der Mittel- und
Hinterbeine deutlich länger [Fig. W31.11].
Innensackstruktur besteht aus einer (spitz zulaufenden) Gabel [Fig. W31.7]. Lebt vornehmlich an Aeonium
canariense. 4,5-8,3mm. Tenerife-Endemit. Aedoeagus [Fig. W31.12].
20 El 2º
segmento abdominal con predominio de estrechas cerdas llamativamente afiladas
hacia la punta [Fig. W31.10]; oniquios
de las patas intermedias y posteriores claramente más largos [Fig. W31.11]. Estructura
del saco interno compuesta por una horquilla (progresivamente afilada) [Fig. W31.7]. Vive
preferentemente sobre Aeonium canariense. 4,5-8,3mm. Endemismo de
Tenerife. Edeago [Fig. W31.12].
.......... Acalles aeonii Wollaston 1864
- 2. Abdominalsegment
mit überwiegend kurzovalen, niemals spitz zulaufenden Borsten [Fig. W31.8]; Klauenglieder deutlich kürzer [Fig. W31.11]. Innensackstruktur besteht aus einer
unterhalb der Spitze verbreiterten
Gabel [Fig. W31.6]. Lebt an Aeonium holochrysum.
4,9-7,8mm. Tenerife-Endemit: östl. Anaga-Gebirge. Aedoeagus [Fig. W31.13].
- El 2º segmento abdominal
con predominio de cerdas cortas, ovales, nunca afiladas hacia la punta [Fig. W31.8]; oniquios
claramente más cortos [Fig. W31.11]. Estructura
del saco interno compuesta por horquilla ensanchada por debajo de su extremo [Fig. W31.6]. Vive
en Aeonium holochrysum. 4,9-7,8mm. Endemismo de Tenerife: macizo de
Anaga oriental. Edeago [Fig. W31.13].
..........
Acalles bodegensis Stüben sp.n."
Beschreibung der Zuchtansatzes:
"F0-Generation"
Um sicherzustellen, dass unbefruchtete Weibchen für
einen sauberen Zuchtansatz zur Verfügung stehen, musste jede Art zunächst
separiert an ihrer jeweiligen Wirtspflanze nachgezüchtet werden.
Acalles bodegensis:
7.1.2004 (Zuchtansatz):
Aeonium holochrysum-Zweige wurden in einen Blumentopf mit Erde und
Blähkügelchen als Drainage, wie sie in der Hydrokultur Verwendung finden,
gepflanzt. Dazu wurden 8 FF und 8 MM von Acalles bodegensis
(FO 9: Bodegas) ins Insektarium gesetzt, die gleich Blatt- und Stängelfraß
aufnahmen. 2 - 5 mm große, oft kreisrunde Löcher wurden ins Blattwerk [Fig. W31.14] und gleich unterhalb der Blattkrone in
die Stängel [Fig. W31.15] sowie vor allem in
einen abgebrochenen, dem Glasboden aufliegenden, 1,5 cm dicken Zweig gefressen
[Fig. W31.16]. Anfänglich kopulierten die Tiere
nach 22.00 Uhr vor allem auf diesem absterbenden Zweig, nur wenige Exemplare
waren auf der Pflanze selbst zu beobachten. [Fig.
W31.17]
23.1.2004 (Rückschlag): Der
Zuchtansatz musste wegen Verpilzung nach zu heftigen, täglichen Besprühungen
wiederholt werden (4 Tiere waren bereits gestorben): Drei Aeonium
holochrysum-Zweige wurden senkrecht in einen Blumentopf mit Sand- und
Blähkügelchen als Drainage gepflanzt. Diesmal wurde außerdem eine 3 cm dicke
Sandschicht um den Blumentopf ins Insektarium eingebracht, auf die direkt ein
1,5 cm dicker Zweig gelegt wurde. Dazu wurden die übrig gebliebenen 7 FF
und 5 MM von Acalles bodegensis gesetzt.
7.2.2004 (weitere Schwierigkeiten): Erneut wurde starke Verpilzung - vor allem in den
Blattkronen - festgestellt. Diese habe ich herausgenommen und das Insektarium
vorderseitig nun mit Gaze verschlossen (Erhöhung der Luftzirkulation). Die
Tiere fraßen danach Löcher direkt in die weitgehend nun blattfreien, dicken
Stängel, in die sie sehr wahrscheinlich auch ihre Eier ablegen bzw. abgelegt
haben. Keine weiteren Beobachtungen!
5.3.2004 (1. Visite): Alle
Elterntiere wurden entnommen. Die Aeonium holochrysum-Stöckchen waren
hohl (von den Larven zerfressen!) und nur noch von der pergamentartigen Rinde
umgeben. Alle Stöckchen wurden jetzt, um im dritten Larvenstadium und der
bevorstehenden Verpuppung jeder Verpilzung vorzubeugen, in ein trockenes
Insektarium überführt und quer auf die Ränder eines Blumentopfes gelegt (um
auch unterseitig stauende Nässe zu vermeiden).
3.4.2004 (2. Visite): Bereits
5 Tage zuvor konnte ein gerade geschlüpftes, tot am Boden liegendes Exemplar
entdeckt werden. Insgesamt wurde die erfolgreiche Nachzucht mit 5 Weibchen
und 14 Männchen abgeschlossen. Alle Exemplare befanden sich noch in ihrer
Puppenwiege und waren fast völlig ausgehärtet. Entweder „klebte“ die
Puppenwiege innenseitig an der pergamentartigen, äußeren Hülle oder befand sich
am Ende im Inneren des Zweiges (im Inneren der splintholzartigen Röhre). Die
Zweige waren völlig vertrocknet und sowohl zwischen der pergamentartigen
Cuticula und der härteren Röhre als auch im Inneren der Röhre selbst von den
Larven völlig zerfressen (raspelholzartige Späne). Die aus diesem Material
gefertigten, sehr harten Puppenwiegen mussten aufgebrochen werden. Die relativ
kleinen Tiere schienen keine Anstalten zu machen, ihre Puppenwiege zu
verlassen. Ferner wurden zu diesem Zeitpunkt nur noch 2 Larven im letzten
Larvenstadium sowie 2 Puppen entdeckt (eine davon wurde in einem
Eppendorfröhrchen durchgezüchtet).
Die
anfänglich sehr schwierige Nachzucht dauerte von der Eiablage bis zur fertigen
Imago gerade einmal 2 ½ -3 Monate. Allerdings lag die Raumtemperatur in der
letzten Woche tagsüber bei 25 °-27 °Celsius, was die Entwicklung und Verpuppung
erheblich beschleunigt haben dürfte. Die Weibchen wurden gleich nach der
Entnahme aus der Puppenwiege von den Männchen getrennt!
Acalles aeonii:
7.1.2004 (Zuchtansatz): Ein Exemplar von Aeonium cuneatum W.
& B. aus Tenerife wurde in einen Blumentopf mit Erde und
Blähkügelchen als Drainage, wie sie in der Hydrokultur Verwendung finden,
gepflanzt [Fig. W31.18]. Dazu wurden 8 FF
und 8 MM von Acalles aeonii (FO 26: Afur) gesetzt, die gleich
Blattfraß aufnahmen. 2 - 5 mm große, oft kreisrunde Löcher bzw. bis zu 3 cm
lange Kavernen wurden meistens unterseitig ins Blattwerk der unteren
Rosettenblätter gefressen [Fig. W31.19]. So
halten sich die Tiere selten auf den oberen kelchartigen Blättern auf, sondern
fressen des Nachts kopfüber an den grundständigen Rosettenblättern [Fig. W31.20]. Tagsüber ziehen sich die Tiere ins
überwiegend harte, abgestorbene Blattwerk zurück und sind hier selbst unter
heftigsten Klopfbewegungen nicht „abzuschütteln“. [Fig.
W31.21]
5.3.2004 (1. Visite): Alle
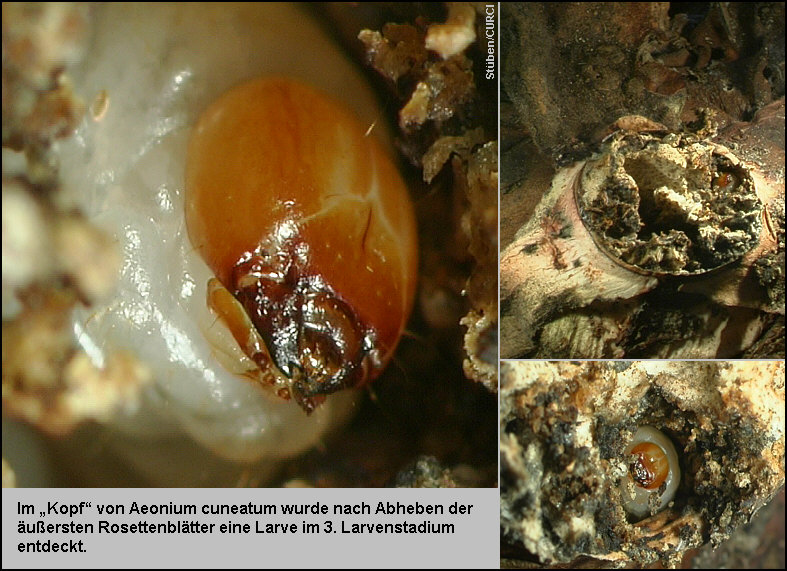
16 eingebrachten Eltern-Exemplare von Acalles aeonii sind tot! Im „Kopf“
von Aeonium cuneatum wurde nach Abheben der äußersten Rosettenblätter
eine Larve im 3. Larvenstadium entdeckt [Fig. W31.22].
3.4.2004 (2. Visite): Bereits
1 Woche zuvor konnten 6 gerade geschlüpfte, tot am Boden liegende Exemplare
entdeckt werden. Zunächst wurden 2 Weibchen und 2 Männchen als Imagines der
Puppenwiege entnommen. Allerdings befanden sich ca. 10 weitere Exemplare noch
im Puppenstadium (unmittelbar vor dem Schlüpfen). Alle Exemplare wurden nach
dem Entfernen der harten, völlig ausgetrockneten Blätter im abgestorbenen
Strunk von Aeonium cuneatum entdeckt. Die Puppenwiegen „kleben“ entweder
an der Innen- oder an der Außenseite des stark durchlöcherten Strunkskeletts.
Die meisten Exemplare wurden noch im Puppenstadium kurz vor dem Schlüpfen
entdeckt und zur Weiterentwicklung - möglichst zusammen mit ihrer Puppenwiege -
in Eppendorfröhrchen überführt. Im unteren Abschnitt des Strunks fanden sich
schließlich nur noch Puppen. Der Strunk wurde dem Pflanztopf schließlich
entnommen und waagerecht auf den sandigen Untergrund des Insektariums gelegt,
um den Puppen eine unbeschadete Weiterentwicklung ohne Staunässe zu
ermöglichen.
Von
der Eiablage bis zur fertigen Imago hat die ganze Nachzucht 3 Monate benötigt.
Ingesamt schlüpften bei dieser Art 6 Männchen und 4 Weibchen, die für
die weitere Kreuzung mit Acalles bodegensis herangezogen wurden. Wo die
Puppen sich zu fertigen Imagines nicht in Eppendorfröhrchen entwickelt haben,
sondern in ihren angestammten Puppenwiegen, wurden sie auch hier wieder diesen
gleich entnommen, um Männchen und Weibchen zu separieren. (Exemplare, die sich
selbst aus der Puppenwiege befreit haben, wurden für die weitere Zucht nicht
herangezogen.)
Beschreibung der
Kreuzung von Acalles aeonii x Acalles bodegensis:
"F1-Generation"
Für die Inselflora der Kanaren sind die adaptiven
Radiationsverläufe der Semperviven gut nachvollziehbar und sowohl in ihrer
intuitiven Erfassung durch frühere Bearbeiter ([Praeger 1932] [Berger 1930])
als auch in ihren verwandtschaftlichen Beziehungen durch die Aufstellung
morphologischer Merkmalswertungen mit daran anschließender Clusteranalyse [Liu
1989] außerordentlich gut erforscht [Lösch 1990]. Die ursprünglichsten
Merkmale der kanarischen Semperviven finden sich bei den Vertretern der
Untergattung Holochrysa, mehr oder weniger unregelmäßig verzweigte
Kleinsträucher, zu denen auch Aeonium undulatum auf Gran Canaria und Aeonium
holochrysum auf den westlichen Inseln gehören. Lems [Lems 1960] sieht in
diesem Formenkreis die Basisgruppe aller kanarischen Arten und Kull [Kull 1982]
nimmt an, "dass Aeonium holochrysum in Gestalt und Ökologie jenen
frühen Aeonium-Arten am nächsten steht, die einst die Kanaren erreichten".
Die Holochrysa-Arten zeichnen sich - im Gegensatz zu den holoxylen Goochia-Arten
und den fast gar nicht verholzten, oligoxylen Canariensia-Arten - durch
den geringen (semixylen) Verholzungsgrad aus. Innerhalb der etwa ein Dutzend Holochrysa-Arten
gehören die genannten Arten von Gran Canaria und Tenerife zu den im Sommer
blatt-abwerfenden Taxa, die einen auffälligen phänologischen Jahresrhythmus
durchlaufen. Mit dem Erwerb der Sommerkahlheit können die Pflanzen auf eine
übermäßige Sukkulenz der Blätter verzichten, was wiederum einen
Bedeutungsrückgang des CAM (Crassulaceae
Acid Metabolism) zur Folge hat.
Die eng miteinander verwandten Wirtspflanzen der Aeonium-Acallen
der westlichen Inseln, nämlich Aeonium
cuneatum (Tenerife), Aeonium canariense (Tenerife), Aeonium palmense (La Palma) und Aeonium
subplanum (La Gomera), gehören in die Aeonium-Sektion Canariensia:
sessile, häufig "kohlkopfartige" Lebensformen außerhalb der
Lorbeerwälder, die an eher trockenen Felsen im Norden der Inseln vorkommen. Auch
hier gibt es keinen Zweifel, dass die Canariensia-Arten von der
ursprünglicheren Basisgruppe der Holochrysa-Arten abgeleitet sind.
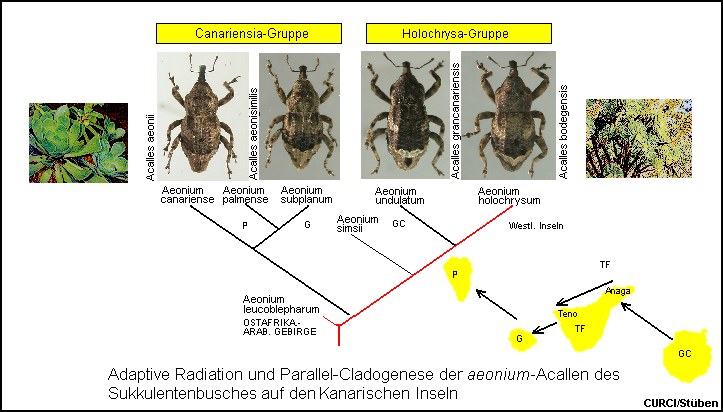
Kontrastiert man die Abstammungsverhältnisse der Aeonium-Pflanzen, an denen Acallen leben, mit den rein morphologisch ermittelten
verwandtschaftlichen Beziehungen unter den Acalles-Arten der aeonium-Gruppe
(Acalles argillosus, der an Kleinia neriifolia lebt, bleibt hier
zunächst unberücksichtigt), fällt eine erstaunliche Deckungsgleichheit auf [Fig. W31.23]. Wir können
jeweils zwei Schwester-Taxa unterscheiden: Acalles grancanariensis und Acalles
bodegensis leben an den eng verwandten Holochrysa-Aeonien, Aeonium
undulatum und Aeonium holochrysum, während die Schwesterarten
Acalles aeonii und Acalles aeonisimilis in den sich besonders nahe
stehenden vier Insel-Vikarianten der Canariensia-Aeonien Aeonium
cuneatum, Aeonium canariense, Aeonium subplanum bzw. Aeonium palmense ihre
Entwicklung durchlaufen. Mit der Abspaltung der dem Boden aufsitzenden Canariensia-Arten
von der Sektion Holochrysa und der Besiedlung der jüngeren
"Inseln" Tenerife-Teno (5-7 Mio.) und La Palma (< 2 Mio.) sowie
der Nachbarinsel La Gomera (15 Mio.) haben sich auch die Aeonium-Acallen
Acalles aeonii und Acalles aeonisimilis von den an Holochrysa
lebenden Arten abgespalten. Die Wirtsspektren und die Biogeographie dieser Taxa
zeigen, dass die Acallen des Sukkulentenbusches tatsächlich in enger
Koevolution eine Parallel-Cladogenese durchlaufen haben und Crassulaceen als
Radiationsplattform für ihre Evolution benutzten. [Stüben 2000k] [Stüben 2000j]
Fazit: Die Nachzucht der beiden Acalles-Arten, Acalles
bodegensis und Acalles aeonii, an der Wirtspflanze Aeonium
holochrysum macht von der Arbeitshypothese Gebrauch, dass für Acalles bodegensis der
Wirtspflanzenwechsel zu Aeonium cuneatum eine
hohe Hürde darstellen könnte, während für Acalles aeonii aufgrund der
"evolutiven Erfahrungen" ein Wirtspflanzenwechsel zu Aeonium
holochrysum vielleicht aufgrund der noch vorhandenen genetischen
Plastizität möglich ist.
Weibchen: Acalles bodegensis x Männchen: Acalles aeonii ("Kleines
Insektarium")
4.4.2004 (Zuchtansatz): 5 Weibchen
von Acalles bodegensis x 6 Männchen von Acalles aeonii werden
an Aeonium holochrysum in das "1. Kleine Insektarium"
eingebracht. Die Einrichtung dieses und der folgenden Insektarien orientiert
sich an den Erfahrungen aus der F0-Zucht: Der Boden wird mit einer ca. 2 cm
dicken, trockenen Sandschicht bedeckt. Das Einbringen eines Blumentopfes mit
Erde und Blähkügelchen als Drainage unterbleibt zukünftig; stattdessen werden
drei bis vier, 1,5 cm dicke und ca. 20-25 cm lange Aeonium holochrysum-Zweige
direkt senkrecht - bzw. gegen die Scheiben des Insektariums gelehnt - auf den
Sandboden gestellt. Nur wenige Blattrosetten bleiben stehen. Sie dienen den
Tieren als Feuchtigkeitsspeicher und Nahrung in den ersten zwei bis drei
Wochen. Auf zusätzliche Besprühungen wird weitgehend verzichtet. [Fig. W31.24]
10.5.2004 (1. Visite): Von
den 11 wurden 9 Tiere wieder gefunden. Alle sind in einer sehr guten
Verfassung, und es zeigt sich, dass die Tiere unter den wenigen Blattrosetten
und den insgesamt trockeneren Habitatbedingungen deutlich aktiver sind. Zwei
weitere Aeonium holochrysum-Zweige werden aufgestellt. Es erfolgte
jedoch zu diesem frühen Zeitpunkt noch keine Suche nach ersten Larven, um teils
schon ausgetrocknetes Entwicklungssubstrat nicht zu beschädigen!
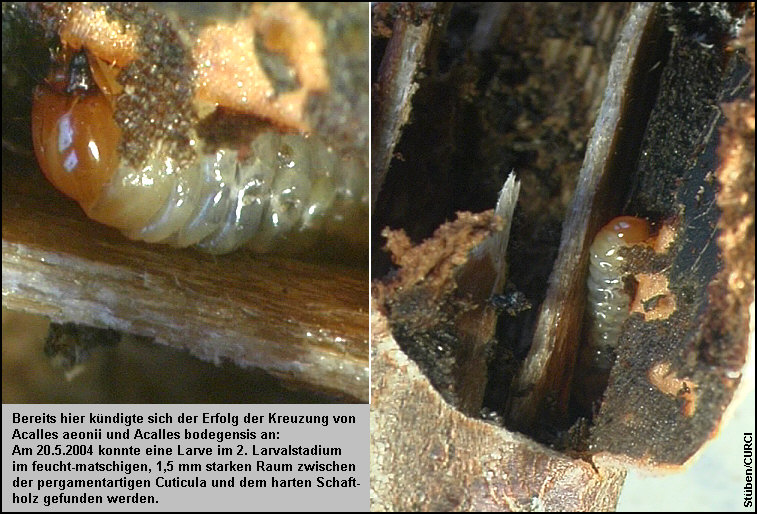
20.5.2004 (2. Visite): In
den letzten Tagen konnten nach 22.00 Uhr an einem der am 10.5.2004
eingebrachten Aeonium holochrysum-Zweigen Tiere bei der Kopulation
beobachtet werden. [Fig. W31.25] Am 20.5. wurde ein im oberen Bereich völlig
ausgetrockneter, in dem dem Boden aufliegenden Bereich feuchter, etwa 10 cm
langer Zweig untersucht. Eine Larve im zweiten Larvalstadium fand sich
im feucht-matschigen, 1,5 mm starken Raum zwischen der pergamentartigen
Cuticula und dem harten Schaftholz. [Fig. W31.26]
Auch diesmal erfolgten keine weiteren
Untersuchungen, um die spärlichen Pflanzenvorräte, die mir nach der jetzt fast
6 Monate dauernden Doppelzucht noch zur Verfügung stand, zu schonen.
25.6.2004 (3. Visite und Einrichtung einer F1-Kontrollgruppe
1): Die 9 Imagines (4 FF/5 MM) der letzten F0-Generation wurden von den Aeonium
holochrysum-Zweigen separiert und in einen neuen Zuchtansatz an zwei
Zweigen in ein neues, "2. Kleines Insektarium" eingebracht (Aufbau
einer F1-Kontrollgruppe 1). Am 9.9.2004 wurden alle 9 Imagines aus diesem
"2. Kleinen Insektarium" abgetötet und präpariert! Ohne Imagines lief
diese Zucht (der F1-Kontrollgruppe 1) dann an zwei, etwas frischeren Zweigen
noch bis zum 11.11.2004 weiter und war sehr erfolgreich! (Siehe 11.11.2004).
16.7.2004 (4. Visite): Eine
sehr kleine (ausgetrocknete) Puppe wird in ihrer Puppenwiege entdeckt!
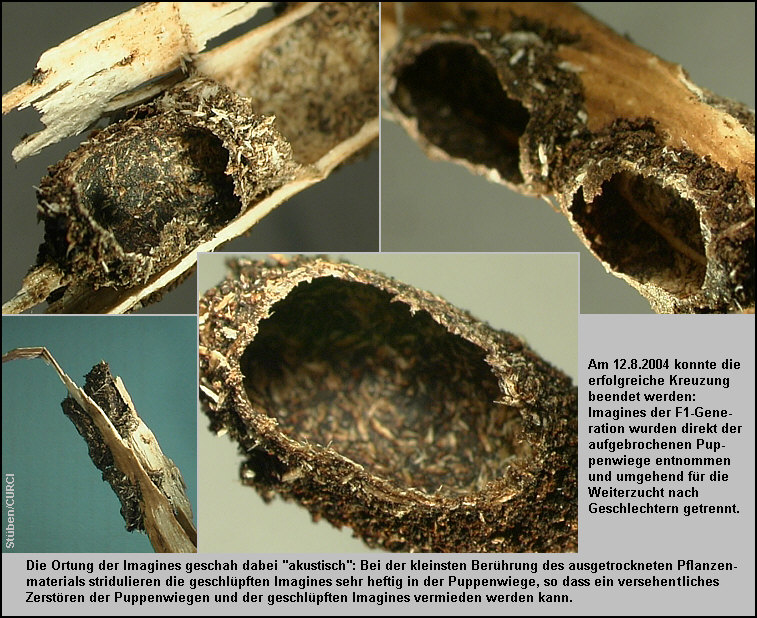
12.8.2004 (Ende der erfolgreichen Kreuzung im "1.
Kleinen Insektarium"): Es wurden insgesamt 7 Exemplare der F1-Generation (3 MM / 4 FF) den jetzt völlig vertrockneten Aeonium holochrysum-Zweigen
entnommen. Sie wurden wie schon die Elterntiere direkt aus den aufgebrochenen
Puppenwiegen gezogen und nach Geschlechtern umgehend separiert. Diese frischen
Imagines befanden sich in ihrer Puppenwiege innerhalb des härteren Schaftes und
- wahrscheinlich aus Platzmangel - in zwei Exemplaren zwischen dem Schaftholz
und der pergamentartigen Cuticula [Fig. W31.27]. Alle Exemplare waren ausgehärtet, aber z.T. noch nicht
ausgefärbt. Ihre Größenunterschiede entsprach in der Bandbreite den
Größenunterschieden der Wildfänge von Acalles bodegensis (1.M=5,4mm;
2.M=4,8mm; 3.M=5,7mm / 1.F=5,5mm; 2.F=6,4mm; 3.F=5,9mm; 4.F=5,2mm).
9.9.2004 (Nachlese): An diesem Tag wurden die letzten
4 Exemplare der F1-Generation der Puppenwiege entnommen. Hier machte ich eine
weitere, sehr interessante Beobachtung: Da die geschlüpften Imagines noch sehr
lange in ihrer Puppenwiege verbleiben, ist neben dem visuellen Aufsuchen der
oft sehr harten Puppenwiegen vor allem die akustische Ortung der Puppenwiegen in
den Zweigen zu empfehlen. Bei der kleinsten Berührung des ausgetrockneten
Pflanzenmaterials stridulieren die geschlüpften Imagines sehr heftig in
der Puppenwiege [Riede & Stüben 2000], so dass ein versehentliches Zerstören
dieser Puppenwiegen und der geschlüpften Imagines vermieden werden kann! [Fig. W31.27] Wieder ergaben
die Größenmessungen der Adultis keine signifikant anderen Werte als bei den Wildfängen
von Acalles bodegensis (4.M=6,0mm; 5.M=5,9mm / 5.F=5,5mm; 6.F=5,5mm). Alle
Exemplare waren ausgehärtet, aber z. T. noch nicht ausgefärbt. Insgesamt wurde
diese erfolgreiche Kreuzzucht im "1. Kleinen Insektarium" mit nur
5 Männchen und 6 Weibchen der F1-Generation abgeschlossen.
11.11.2004 (Ende der erfolgreichen Kreuzung im "2.
Kleinen Insektarium" der F1-Kontrollgruppe 1): Die an zwei frischen Zweigen am 25.6.2004
aufgenommene Kreuzung der bereits am 9.9.2004 abgetöteten 9 Eltern (4 FF/5 MM) wurde am 11.11.2004 beendet. Diese Kontrollgruppe bestand aus 21
Männchen, 23 Weibchen und 6 toten bzw. verletzten Exemplaren. Nur 12 MM
und 12 FF wurden für die Weiterzucht benötigt, die anderen Exemplare wurden
abgetötet und präpariert!
Alle Tiere mussten aus ihrer steinharten Puppenwiege befreit
werden. In einem der etwa 25 cm langen Aeonium holochrysum-Zweige
befanden sich auf engstem Raum (befallener Abschnitt: ca. 5 - 8 cm lang) allein 27 zum Teil
sehr kleine Exemplare. Dies ist ebenfalls eine sehr wichtige Beobachtung, die
meine schon oft geäußerte These bestätigt, dass die Größe aller
Cryptorhynchinae im Wesentlichen vom Raumangebot des
Entwicklungssubstrates - und damit auch
der Nahrungsmenge pro Larve - bestimmt wird. Daher darf Größenangaben bei
Cryptorhynchinae grundsätzlich keine Bedeutung beigemessen werden!
Männchen: Acalles bodegensis x Weibchen: Acalles aeonii ("Großes
Insektarium")
4.4.2004 (Zuchtansatz): 14
Männchen von Acalles bodegensis x 4 Weibchen von Acalles
aeonii werden an Aeonium holochrysum in das "Große
Insektarium" eingebracht.
10.5.2004 (1. Visite): Von
den 18 wurden 15 Tiere zunächst wieder gefunden. Wieder waren alle Tiere in
einer sehr guten Verfassung. Zwei weitere Aeonium holochrysum-Zweige
wurden eingebracht. Auch hier unterblieb zu diesem frühen Zeitpunkt die Suche
nach ersten Larven, um das Entwicklungssubstrat zu schonen.
25.6.2004 (2. Visite und Einrichtung einer F1-Kontrollgruppe
2): Von
den ursprünglich 18 Tieren wurden alle Exemplare wieder gefunden, davon
wiederum waren 10 Exemplare verstorben. 2F / 6MM der letzten
F0-Generation wurden von den Aeonium holochrysum-Zweigen separiert und
in einen neuen Zuchtansatz in ein "2. Kleines Insektarium"
eingebracht (Aufbau einer F1-Kontrollgruppe 2). Am 9.9.2004 wurden alle
8 Adultis abgetötet und präpariert! Ohne Adultis lief die Zucht (der
F1-Kontrollgruppe 2) an drei noch etwas frischeren Zweigen bis zum 11.11.2004
weiter (siehe dort)!
16.7.2004 (3. Visite): Im
"Großen Insektarium" wurde an zwei Zweigen jeweils eine Larve im 3.
Stadium entdeckt!
12.8.2004 (Ende der erfolgreichen Kreuzung im "Großen
Insektarium"):
Es wurden insgesamt 5 Exemplare
(2 MM / 3 FF) den jetzt völlig
vertrockneten Aeonium holochrysum-Zweigen entnommen. Sie wurden wie schon die Elterntiere direkt aus den
Puppenwiegen „gebrochen“. Diese befanden sich alle innerhalb des härteren
Schafts (Größe: 1. M=6,1mm; 2. M=5,4mm / 1. F=7,8mm; 2. F=6,3mm; 3. F=5,7mm).
Alle Exemplare waren ausgehärtet, aber z. T. noch nicht ausgefärbt. (Die im
Durchschnitt deutlich größeren Tiere ergeben sich aus den viel dickeren Aeonium
holochrysum-Zweigen dieses Zuchtansatzes!)
9.9.2004 (Nachlese): An diesem Tag wurden die letzten
8 Exemplare der F1-Generation der Puppenwiege entnommen: 3. M=5,4mm; 4. M=6,9mm
5. M=7,0mm; 6. M=4,5mm / 4. F=6,6mm; 5. F=6,0mm; 6. F=5,7mm; 7. F=5,6mm. Alle
Exemplare waren ausgehärtet, aber z. T. noch nicht ausgefärbt. Insgesamt wurde
diese erfolgreiche Kreuzzucht im "Großem Insektarium" mit 6
Männchen und 7 Weibchen der F1-Generation abgeschlossen.
11.11.2004 (Ende der erfolgreichen Kreuzung im "2.
Kleinen Insektarium" der F1-Kontrollgruppe 2): Die an drei frischen Zweigen am 25.6.2004
aufgenommene Kreuzung der bereits am 9.9.2004 abgetöteten 8 Eltern (2 FF/6 MM) wurde am 11.11.2004 beendet. Diese Kontrollgruppe 2 ergab
zunächst 1 Männchen, 4 Weibchen und 4 Puppen. Aus den vier Puppen entwickelten sich am 25.11.2004 4 Imagines, 2 Männchen und
2 Weibchen. Insgesamt entstammen dieser F1-Kontrollgruppe 2 also 3
Männchen und 6 Weibchen, die für die Weiterzucht Verwendung fanden. Alle
Tiere mussten aus ihrer steinharten Puppenwiege befreit werden.
Die
erfolgreiche Kreuzung hat also in den beiden Hauptinsektarien
("Kleines" und "Großes Insektarium") und in den beiden
Kontrollgruppen etwa 4 Monate in Anspruch genommen. Das erscheint deutlich
länger als in der F0-Generation, darf aber nicht überbewertet werden, da in der
Zwischenzeit doch allmählich die Fraß- und Entwicklungspflanze Aeonium holochrysum knapp
wurde und natürlich die Imagines nicht immer gleich der Puppenwiege
entnommen werden konnten, in der sie sicherlich einige Wochen verharren können,
ohne Anstalten zu machen, sich selbst "zu
befreien"!
Was
lässt sich über die morphologischen Unterschiede der Individuen der
F1-Generation feststellen? Dazu wurden die am 11.11.2004 "geschlüpften" und abgetöteten Imagines
der F1-Kontrollgruppe 1 näher untersucht. Unter den 25 Exemplaren fanden
sich sowohl Tiere mit langen, schlanken als
auch kurzen, dicken Borsten unter dem 2. Abdominalsegment. Bei den
Klauengliedern der Mittel- und Hinterbeine konnten in der Länge keine
signifikanten Abweichungen mehr festgestellt werden, während die Gabel der
Innsackstruktur des Aedoeagus in allen Fällen (wie bei Acalles aeonii)
spitz zuläuft. In der tiefen und grubenartigen Punktur der Elytrenflanken sowie
den kontrastreichen Hell-dunkel-Zeichnungen der Elytren entsprachen die
Exemplare dieser Kontrollgruppe 1 wiederum ganz dem Aussehen von Acalles
bodegensis. Die Durchschnittsgröße der Imagines bewegte sich eher im
unteren Bereich und lag damit im Größenspektrum von Acalles bodegensis.
Doch erlauben diese letzten Angaben (siehe oben) keine weiterführenden
Schlussfolgerungen, da - wie bei allen Cryptorhynchinae - die Körpergröße im
Wesentlichen vom Raumangebot des Entwicklungssubstrates - und damit auch der Nahrungsmenge
pro Larve - bestimmt wird. Es dürfte einleuchten, dass die schlanken Zweige von
Aeonium holochrysum den Tieren ein
deutlich geringeres Raum- und Nahrungsangebot bereitstellen (vor allem unter
den Bedingungen der doch relativ kleinräumigen Insektarien) als die sehr dicken
Strünke der "Kohlkopf-förmigen" Aeonium cuneatum-Pflanzen. Ingesamt handelt es sich bei diesen Untersuchungen, die
nicht im Focus meines Interesses standen, natürlich nur um eine
Stichprobe.
Die
entscheidende Frage aber ist, ob an dieser Stelle die genetische Differenz noch
reversibel ist, also vielleicht "fruchtbare Hybride"
entstanden sind? Diese Frage kann nur durch eine Weiterzucht beantwortet
werden. Wird sie bejaht werden müssen, sollte man nicht mehr von zwei
"Arten" sprechen. Die "Arten" wären in diesem Fall eben nicht
reproduktiv isoliert! (Schon die Annahme, dass vielleicht in Zukunft die Arten
im Anaga-Gebirge auf Tenerife weiter divergieren könnten - für manche Autoren
Anlass von "Unterarten" zu sprechen - gehört hingegen ins Reich der
Spekulationen; dazu [Stüben 2003: 215]).
Beschreibung der
Weiterzucht von Acalles aeonii x Acalles bodegensis: "F2-Generation"
Die Einführung zweier Kontrollgruppen (und damit die
Erreichung einer zeitlichen Verzögerung um 3 Monate) war notwendig geworden,
weil ich auf diese Weise mit der F2-Zucht einmal im August, also im Spätsommer,
und ein weiteres Mal im November, also im Herbst 2004, beginnen konnte. Die
Länge der Tage, die Intensität der Sonneneinstrahlung und die Zahl der Sonnenstunden
pro Tag schwanken doch erheblich zwischen den Nordhängen des Anaga-Gebirges auf
Tenerife und dem beheizten Wintergarten des CURCULIO-Institutes am Niederrhein
(Germany). Außerdem konnte ich nicht davon ausgehen, dass sich die Tiere bei
einer relativ konstanten Zimmertemperatur von 24 °C und einer relativen
Luftfeuchtigkeit von 45% - 55% tatsächlich zu allen Jahreszeiten reproduzieren
würden.
Meine
zur Neige gehenden Bestände an Aeonium holochrysum zwangen mich darüber
hinaus zu einer ganz ungewöhnlichen Bitte um Hilfe von den Kanarischen Inseln,
und ich war froh, dass rechtzeitig noch Aeonium holochrysum-Zweige aus
dem eigenen Anbau meines Kollegen Antonio Machado (La Laguna / Tenerife)
eintrafen. Ihm sei an dieser Stelle herzlich gedankt! Selbstverständlich wurden
die Zweige nach Fraß- und möglichen Eiablagespuren unter dem Mikroskop
genauestens untersucht. Ein Befall mit Eiern oder Larven lag nicht vor und so
konnte die zweifellos aufwendige und nicht ganz leichte Zucht also im August
bzw. November 2004 in ihre vorläufig letzte Phase gehen. [Fig. W31.28]
Nachzucht der F2-Generation im "Roten
Insektarium"
13.8.2004 (Beginn der F2-Zucht): 3 MM (Kleines Insektarium) x 3 FF (Großes Insektarium).
An 2 x 1,5 cm dicken Aeonium holochrysum-Zweige aus eigenem Anbau und an
2 x 3 cm dicken Aeonium holochrysum-Zweige aus der „Lieferung Machado“
wurden zunächst 6 Tiere der F1-Generation gesetzt. Am 9.9.2004 wurden
weitere 2 MM (Kleines Insektarium) x 4 FF (Großes Insektarium)
dazugesetzt. Ein weiterer dicker Aeonium holochrysum-Zweige aus der
„Lieferung Machado“ wurde ebenfalls eingebracht. Insgesamt: 5 MM
(Kleines Insektarium) x 7 FF (Großes Insektarium).
20.12.2004 (3. Visite): Alle
F1-Elterntiere wurden wiedergefunden, entfernt, abgetötet und präpariert. Das Rote Insektarium jetzt nur noch mit 5 kurzen
Zweigen (in einem Zweig wurde eine weitere Puppe entdeckt).
22.1.2005 (Ende der erfolgreichen F2-Zucht im
Roten Insektarium): Weitere 3FF und 2MM wurden ihren
Puppenwiegen direkt entnommen [Fig. W31.29]
sowie 1 Larve (3. Stadium) und 2 Puppen [Fig. W31.30]
in Alkohol überführt. Insgesamt wurden 3MM und 6FF der
F2-Generation im Roten Insektarium
nachgezüchtet und zur Weiterzucht in ein F3-Insektarium überführt.
Die Folgenden F2-Nachzuchten werden nicht mehr ausführlich geschildert. Es sollen hier nur kurz die wichtigsten Beobachtungen und Ergebnisse festgehalten werden.
Nachzucht der F2-Generation im "Blauen
Insektarium"
13.8.2004: 6FF (Kleines Insektarium) x 6MM
(Großes Insektarium). Nur an einem schlanken Aeonium holochrysum-Zweig
(aus eigenem Anbau) konnten zunächst Eiablageeinstiche entdeckt werden [Fig. W31.31]. Dieser Zweig wurde erst am 9.9.2004
eingebracht, enthielt aber bereit am 29.11.2004 eine Larve im letzten Stadium,
die sich am 10.12.2004 bereits verpuppt hatte. In diesen kaum verholzten,
grünen Zweigen läuft die Entwicklung deutlich schneller ab als in den kräftigen
Ästen aus der „Lieferung Machado“. Solche starken, schon bei der Anlieferung
weitgehend verholzten „Stämme“ von Aeonium holochrysum wurden ganz
offensichtlich nicht angenommen und lieferten auch keine weiteren Nachzuchten.
[Fig. W31.28]
Insgesamt
konnte nur 1 F2-Puppe im überwiegend mit stark verholzten Aeonium
holochrysum -„Stämmen“ bestückten Blauen
Insektarium nachgezüchtet werden, die sich jedoch außerhalb der
Puppenwiege nicht mehr zur fertigen Imago weiterentwickeln konnte.
Nachzucht der F2-Generation im "Grauen
Insektarium"
11.11.2004:
Beginn der F2-Zucht. Ganz anders
fiel die Nachzucht von 12 (der ursprünglich 21) MM und 12 (der
ursprünglich 23) FF aus der „Kontrollgruppe 1 des Kleinen Insektariums“
aus (siehe oben). Die fünf, ebenfalls aus der „Lieferung Machado“ stammenden Aeonium
holochrysum-Zweige (ca. 20 cm lang und 2-3 cm dick) waren noch stellenweise
grün und deutlich weniger verholzt. Am 22.1.2005 fanden sich bereits zahlreiche
Larven im letzten Larvenstadium. Ebenso wurden bei einer weiteren Kontrolle am
13.2.2005 2 weitere Larven im letzten Larvenstadium in bereits vorgefertigten
Puppenwiegen nachgewiesen.
Zweige, die befallen sind, sind zu diesem Zeitpunkt leicht an der
„aufgeblähten“, pergamentartigen Cuticula zu erkennen. Sie fühlen sich weich an
und lassen sich leicht eindrücken. Entscheidend ist, dass ab diesem Zeitpunkt
die Zweige austrocknen müssen und daher auf keinen Fall mehr besprüht werden
dürfen, denn die Puppen benötigen ein absolut trockenes, sehr hartes Medium, um
sich in ihrer Puppenwiege einrichten zu können. Am 18.3. und 31.3.2005 konnten
diesen Zweigen im Grauen Insektarium insgesamt
8 MM und 4 FF der F2-Generation entnommen werden und zur
Weiterzucht in ein F3-Insektarium überführt werden.
Weitere Nachzuchten
aus dem Roten Insektarium und dem Grauen Insektarium
Zwischen dem 22.1. - 31.3.2005 wurden insgesamt
21 Exemplare (11 MM und 10 FF) aus dem Roten Insektarium und dem Grauen
Insektarium für die F3-Weiterzucht in ein letztes gemeinsames
Insektarium überführt. Am 23.4.2005 konnten von den insgesamt 21
eingebrachten Exemplaren noch 15 sehr vitale Tiere nachgewiesen werden.
Auch bei den verstorbenen Tieren (die Luftfeuchtigkeit war während der
Wintermonate sehr stark abgefallen!) konnten keine Anomalien bzw. Missbildungen
beobachtet werden. Ende Mai 2005 wurde eine erste Puppe nachgewiesen. In
derselben Puppenwiege wurde am 8.6. 2005 das erste noch immature F3-Exemplar
freigelegt.
10.7.2005: Die
F3-Zucht erbrachte insgesamt 3 MM + 2 FF. 1 Tier war beim
Aufschneiden verletzt worden. Die Tiere mussten mangels Fraßpflanze abgetötet
werden! Damit endete die 1 ½-jährige Nachzucht. Rechnet man die F0-Generationen
von Acalles aeonii und Acalles bodegensis syn nov. dazu, waren es
genau 4 Generationen!
3. Ergebnisse und Diskussion
Quod erat demonstrandum: Acalles
aeonii Wollaston 1864
= Acalles bodegensis Stüben 2000
Dieses Ergebnis des Kreuzungsexperiments hatte ich in dieser Eindeutigkeit nicht erwartet. Die Art Acalles bodegensis Stüben 2000 ist damit nachweislich ein jüngeres Synonym von Acalles aeonii Wollaston 1864!
Die morphologischen Unterschiede von Individuen "innerhalb" einer Art (besser: Art-Population) sollten kleiner sein als zwischen den Arten (Art-Populationen). "Messen" lassen sie sich in den seltensten Fällen und wo dies dennoch möglich ist, erlaubt weder die genetische noch die morphologische Distanzmatrix a priori eine Entscheidung darüber, ob es sich um eine oder zwei Arten handelt. Wir müssen uns - wohl oder übel - jenseits von Kreuzungsexperimenten damit abfinden, dass es hier keine "objektiven" Abgrenzungskriterien gibt. Wo in der Praxis Kreuzungsexperimente unterbleiben, gehören ein oft äußergewöhnlicher Spürsinn und eine große Erfahrung dazu, "um das Ausmaß der möglichen intraspezifischen Variabilität einschätzen zu können" [Wägele 2001].
Um so wichtiger ist
es, von Zeit zu Zeit solche Kreuzungsexperimente auch tatsächlich
durchzuführen, um subjektive morphologische Differenzierungserfahrungen neu zu
justieren bzw. an der "Realität" scheitern zu lassen!
So reichen in meiner
Erstbeschreibung von Acalles bodegensis offensichtlich die minimalen
Unterschiede bei der Innensackstruktur des Aedoeagus,
dem 2. Abdominalsegment und der Länge der Klauenglieder nicht aus, um diese Art
von Acalles aeonii tatsächlich "abspalten" zu können (siehe
oben). Auch die unterschiedliche Pflanzenbindung an den verschiedenen Fundorten
- hier Aeonium holochrysum, dort Aeonium cuneatum - ist nicht mehr als ein Artefakt, letztlich eine durch
menschliche Wahrnehmung geschaffene „Illusion“.
Das
heißt nun nicht, dass solche Unterschiede nicht vorhanden wären, dass sie nicht
irgendwann in einen Prozess der Spezifikation einmünden. Doch wir können dieses Spezifikationsereignis weder
voraussagen noch durch eine bloße "taxonomische" Entscheidung für
eine neue Art oder Unterart präjudizieren! Die Evolution lässt sich nicht in
die Karten schauen, schon gar nichts „vorschreiben“, Systematiker bleiben
"Konstrukteure", und die Taxonomie sollte kein Basar persönlicher
Einschätzungen und subjektiver Rechthabereien oder Eitelkeiten sein.
Finden
wir uns also damit ab, dass es Übergangsfelder gibt, in denen wir Populationen
als "Rassen" oder "Rassenkreise" von Arten betrachten. Das
ist allemal besser als vollendete Tatsachen vorzuspiegeln, wo es
in der Evolution (bisher) keine gegeben hat! Kreuzungsexperimente können uns
von Zeit zu Zeit dabei weiterhelfen - die selbst gestrickte
"Ausrede", solche Experimente seien zu schwierig oder gar
undurchführbar, aber gewiss nicht!
4. Zum Schluss: Eine Gesprächsnotiz
Der Autor beim
Verlassen des Sequenzierungslabors (mit resignativem Blick):
„Sie wollen mir bei der Artdifferenzierung also
nicht mit Rat und Tat zur Seite stehen, wo ich alles so schön über Jahre hinweg
in Alkohol konserviert habe?“
Molekularbiologe
(mit verständnisvollem Blick):
„Von „wollen“ kann wirklich nicht die Rede sein. Seien
sie doch kein Narr und machen die gleichen methodologischen Fehler wie unsere
Art-Sequenzierungsanalytiker, die besser den Alkohol saufen sollten, in dem Sie
ihre vermeintlichen Arten für ihre Analysen konserviert haben. Beschäftigen Sie
sich endlich mal mit „reproduktiver Isolation“, mit dem biologischen Artbegriff
und mit Sex. Überlassen Sie es ihren Acalles-Arten, ihnen zu helfen!
Züchten Sie! Das ist billiger und schafft Klarheit – oft endgültige Klarheit.
Aber so etwas ist heute leider aus der Mode gekommen.
Machen Sie es gut!
5. Literatur
Berger A. (1930): Crassulaceae. - In: A. Engler, K. Prantl (Hrsg.): Die
natürlichen Pflanzenfamilien. 2. Aufl., 352-386, Leipzig.
ICZN (2000): Internationale Regeln für die zoologische Nomenklatur
(offizieller deutscher Text, ausgearbeitet von O. Kraus), - Abhandlungen des
Naturwissenschaftlichen Vereins in Hamburg (NF), 34: 232 S., Goecke
& Evers: Keltern-Weiler.
Kull U. (1982): Artbildung durch geographische Isolation bei Pflanzen -
die Gattung Aeonium auf Teneriffa. - Natur und Museum 112: 33-40.
Lems K. (1960): Botanical notes on the Canary
Islands. II. The evolution of plant forms in the islands: Aeonium. -
Ecology 41: 1-17.
Liu H.-Y. (1989): Systematics of Aeonium (Crassulaceae). - Natl. Museum
of Nat. Sci. Taiwan, Special Publicat. 3, 102 S.
Lösch R. (1990): Funktionelle Voraussetzungen der adaptiven
Nischenbesetzung in der Evolution der makaronesischen Semperviven. -
Dissertationes Botanicae, Bd. 146, 482 S., Berlin / Stuttgart.
Mayr E. (1942): Systematics and the origin of
species. Columbia University Press, New York.
Mayr E. (1969): Principles of Systematic Zoology. Mac-Graw-Hill Book
Co., New York.
Praeger R.L. (1932): An account on the Sempervivum group, 265 S.,
London. (Reprint 1967).
Riede K. & Stüben, P.E. (2000): "Die musikalischen
Acallen": Beobachtungen zur Stridulation bei den Cryptorhynchinae von den
Kanaren. - Cryptorhynchinae-Studie 13 - in: Stüben, P.E. (2000b), SNUDEBILLER 1
(CD ROM): 307-317, Mönchengladbach.
Stüben, P.E. (2000b): (Ed.), Die Cryptorhynchinae der Kanarischen
Inseln. Systematik, Faunistik, Ökologie und Biologie. - SNUDEBILLER 1
(CD ROM); mit 910 Farbfotos, 266 REM-Aufnahmen, 118 Verbreitungskarten, 18 Ton-
u. 1 Video-Aufnahme, 1. Edition, CURCULIO-Institute, D-Mönchengladbach: 413 pp.
(662MB).
Stüben P.E. (2000e): Die Arten des Genus Acalles von den
Kanarischen Inseln. - Cryptorhynchinae-Studie 5 - in: Stüben, P.E. (2000b),
SNUDEBILLER 1 (CD ROM): 22-98, Mönchengladbach.
Stüben P.E. (2000j): Biogeographie und Evolution der kanarischen
Cryptorhynchinae. - Cryptorhynchinae-Studie 10 - in: Stüben, P.E. (2000b),
SNUDEBILLER 1 (CD ROM): 293-306, Mönchengladbach.
Stüben P.E. (2000k): Phylogenie der endemischen Taxa des Genus Acalles
von den Kanarischen Inseln. - Cryptorhynchinae-Studie 12 - in: Stüben, P.E.
(2000b), SNUDEBILLER 1 (CD ROM): 287-292, Mönchengladbach.
Stüben, P. E. (2003): Beschreibung neuer Cryptorhynchinae aus Spanien,
Italien und Marokko - mit einem neuen Onyxacalles- und Echinodera-Schlüssel.
- SNUDEBILLER 4, Studies on taxonomy, biology and ecology of
Curculionoidea, Mönchengladbach: CURCULIO-Institute, 186-224.
Stüben P.E., Behne, L. & Bahr, F. (2003): Analytischer Katalog der
westpaläarktischen Cryptorhynchinae / Analytical Catalogue of Westpalearctic
Cryptorhynchinae. Teil/Part 2: Acalles, Acallocrates (Col.:
Curculionidae: Cryptorhynchinae). - SNUDEBILLER 4, Studies on taxonomy,
biology and ecology of Curculionoidea, Mönchengladbach: CURCULIO-Institute, 11-100.
Wägele J-W. (2001): Grundlagen der Phylogenetischen Systematik. 320 S.,
München, 2. Auflage.
Wollaston, T. V.
(1864): Catalogue
of the Coleopterous insects of the Canaries in the Collection of the British
Museum: 283-298; London.
Adresse des Autors
Dr. Peter E. Stüben
CURCULIO-Institut
Hauweg 62
D-41066 Mönchengladbach
E-Mail: P.Stueben@t-online.de
* Vortrag gehalten am 5. März 2005 anlässlich der Jahrestagung der
„Gemeinschaft für Coleopterologie“ (COLEO, Germany)
Eine Kurzfassung dieses Beitrags findet sich auch in: Stüben, P.E. &
Christoph Germann (2005): Neue Erkenntnisse zur Taxonomie, Biologie und
Ökologie der Cryptorhynchinae von den Makaronesischen Inseln. 1. Beitrag:
Kanaren / Tenerife (Coleoptera: Curculionidae: Cryptorhynchinae) - SNUDEBILLER 6,
Studies on taxonomy, biology and ecology of Curculionoidea, Mönchengladbach:
CURCULIO-Institute, 84-113.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}