Weevil News
|
http://www.curci.de/Inhalt.html |
No.
19 |
13
pp. |
30.
Juni 2004 |
ISSN 1615-3472
|
|
Stüben
P.E. (2004g):
Zur Biologie von Acalles poneli Stüben
2000 (Coleoptera: Curculionidae: Cryptorhynchinae)-
Weevil News: http://www.curci.de/Inhalt.html, No. 19:
13 pp., CURCULIO-Institute: Mönchengladbach. (ISSN
1615-3472). |
|||||
Zur
Biologie von Acalles poneli Stüben 2000
(Coleoptera: Curculionidae: Cryptorhynchinae)
von
Peter E. Stüben, Mönchengladbach
mit 35 Farbtafeln, 13 REM-Aufnahmen, 1
Verbreitungskarte
[Stüben]
Abstract
On the Biology of Acalles poneli Stüben 2000
(Coleoptera: Curculionidae: Cryptorhynchinae); with 35 coloured tables, 13 SEM
photos and 1 distribution map; including a summary in English language.
The
host plant relationships of Acalles poneli Stüben 2000 from Tenerife
(Teno Mts) are studied in detail. Not Euphorbia obtusifolia Poir., as
supposed up to now, but Euphorbia balsamifera Ait. and Euphorbia
atropurpurea (Brouss.) W. & B. are the host plants of Acalles poneli.
The adult weevils have to overcome a high barrier: the insect-toxic latex of
Euphorbiaceae (‘avoidance strategy’). The successful breeding of Acalles
poneli on Euphorbia atropurpurea, host plant of ‘first choice’ is
presented here, and the different environmental parameters effective during
larval stages are compared with habitat conditions on Tenerife. The result: Not
only the presence of the host plant Euphorbia atropurpurea is a crucial
pre-condition, but also the rapid change from extreme wet to extreme dry habitat
structures (‘switch-habitats’) during metamorphosis is important for the
moment of egg-deposition in Acalles poneli! Finally eggs and larvae of Acalles
poneli are described for the first time.
Keywords
Coleoptera,
Curculionidae, Cryptorhynchinae, Acalles poneli, Euphorbia
atropurpurea, Euphorbia balsamifera, biology, ecology, breeding,
host plant, first description of larva, eggs, Spain, Canary Islands,
Tenerife.
Results
and discussion:
The ‘switch-habitat thesis’
1.
The breeding of Acalles poneli Stüben 2000 [Fig.
1][Fig. 2]
presented here confirms the previous experiences with the breeding of
Cryptorhynchinae: The development from egg-deposition until appearance of the
imago lasts about 3 months. [Fig.
31E] The first 4-6 weeks between January, 7th and May, 9th
2004, there was no deposition of eggs on the still latex-containing isolated
twigs of Euphorbia atropurpurea in the insectaries of CURCULIO
Institute. [Fig. 18E]
The compounds of the latex of Euphorbiaceae toxic for insects are well-known
for a long time. [Hegnauer 1966: 106 -140] [Fig.
7cE] A successful larval development will only start when the latex
supply has stopped. [Sprick & Stüben 2000] The
process of desiccation is passing off very slowly below the cuticula of Euphorbia
atropurpurea, and evaporation is minimized: The twigs cut off on Tenerife
contained partly latex below the leaf rosettes still 6 months later. [Fig.
19E]
2.
How has Acalles poneli adapted to these very difficult
environmental conditions? Observations and the situation of detection of Acalles
poneli and its host plant Euphorbia atropurpurea in the extreme
west of Teno Mountains (Tenerife) are important here. [Fig.
DaPON] The plants colonized by this Cryptorhynchinae grow at the
foot of a rock face belonging to a spring water slope of the Barranco del
Carrizal. The slope was very wet in winter 2003/2004.
[Fig. 14E]
At this time the Barranco del Carrizal still contains water, but is completely
running dry during the hot summer months. Then, the trade wind clouds coming
from the north reach the western corner of Teno massif only for a few days.
All these factors may explain the high percentage of dead Euphorbia
atropurpurea-shrubs on the spring water slope at least wet during winter
month: The hard change
from extreme wet to extreme dry habitat conditions is responsible for a
definite susceptible population of stressed plants and an ideal area for
endophytic life-forms. [Fig.
13E] [Fig. 15E]
In winter 2003/2004, we really didn’t find any larvae on the vigorous Euphorbia
atropupurea-shrubs containing a large amount of latex. Not until the
complete drying off of the rocky slope numerous plants will die; a process
that must begin very early in the year (April?): Even detached twigs keep the
insect-toxic latex below their cinnamon-coloured bark. Therefore, my main
thesis is:
3. At the ‘locus typicus’, the north of Teno Mts., Acalles poneli also lives on Euphorbia balsamifera. [Fig. 9E] The occurrence of the real host plant Euphorbia atropurpurea is necessary, but it is not a sufficient pre-condition for the egg-deposition and the subsequent development into the imago stage: Not only the occurrence of the host plant Euphorbia atropurpurea is crucial for the development of Acalles poneli, but the rapid change from extreme wet to extreme dry habitat structures (‘switch-habitats’). This fits in with two observations: A. The spring water slope mentioned above on a steep rock face extends eastwards where a population of Euphorbia atropurpurea is growing under drier conditions for the whole year. Here only very few specimens of Acalles poneli were found (relation about 10:1). B. The above-mentioned breeding has also shown: “Sufficient humidity” during the two first larval stages is the most decisive parameter for a successful development of Acalles larvae; here between the parchment-like bark and the hard sapwood-like stem. [Fig. 27E] Only just the larvae of the last, third stage leave the ‚squashy’, dark brown to black ‘ intervening space’ and eat themselves through the hard stem into the medulla of the Euphorbia atropurpurea-twigs [Fig. 28E], that has dried out now to a great extent; here they pupate afterwards. [Fig. 29E] During this phase larvae and pupae absolutely need extreme dry habitat conditions!
4. As a rule, the egg-deposition of our flightless Cryptorhynchinae is not bound to a definite season, as could be shown by breeding Acalles poneli. On the Canary Islands, the seasonal fluctuation of temperatures is well-balanced. There is no month without precipitation in the north-eastern Las Mercedes-forest of Anaga Mountains (more than 150 mm per month from December to February). Wherever is a chance, for example in case of a stress-caused dying of the plants (as shown here) or by mechanical effects (as in partial wind- or scree break), the animals will start immediately(!) with egg-deposition after interruption (decomposition) of the toxic plant metabolites. [Fig. 20E] That they prefer a phase for the beginning of development being within a period of humidity and changing at the latest with the emergence of the third larval stage into an extreme dry period may be supposed. To start egg-deposition at the beginning of a humid period in a latex-free segment of the plant is forbidden by the risk of wet: The cuticula of many Euphorbiaceae stops evaporation to a large extent. (During the two first larval stages, even very low spraying resulted in an early fungus infection of the larvae below the parchment-like cuticula. [Fig. 26E]) On the other hand an egg-deposition on the peak of a dry phase is ruled out by the same reasons. But never before, Acalles species were observed to deposit their eggs to dry or dead wood! (Even if this is claimed by the complete misjudging of the indispensable complex biotic and abiotic pre-conditions of a successful metamorphosis in Cryptorhynchinae. [Kippenberg 1983:162] [Koch 1992: 301]).
Zusammenfassung
Ausführlich werden die
Wirtspflanzenbindungen von Acalles poneli Stüben 2000 auf Tenerife (Teno
Mts.) beschrieben. Nicht Euphorbia obtusifolia
Poir., wie bisher angenommen, sondern Euphorbia balsamifera Ait. und Euphorbia
atropurpurea (Brouss.) W.&B.
sind die Fraß- und Entwicklungspflanzen von Acalles poneli. Dabei gilt
es für die Imagines eine hohe Hürde zu nehmen: die insektentoxisch wirkenden
Milchsäfte der Euphorbiaceae („Vermeidungsstrategie“). Die erfolgreiche
Zucht von Acalles poneli an Euphorbia
atropurpurea,
der Wirtspflanze der „ersten Wahl“, wird vorgestellt, und die
unterschiedlichen Umweltparameter während der verschiedenen Larvalstadien
werden mit den tatsächlichen Habitatstrukturen auf Tenerife verglichen. Das
Ergebnis: Nicht so sehr das Vorhandensein
der Wirtspflanze Euphorbia atropurpurea an sich ist entscheidend, sondern
die Möglichkeit des raschen Wechsels von extrem feuchten zu extrem
trockenen Habitatstrukturen („Wechselhabitate“) während der Metamorphose
sind für Acalles poneli und für den Zeitpunkt der Eiablage
ausschlaggebend!
Abschließend
werden erstmalig die Eier und Larven von Acalles poneli beschrieben.
Philippe Ponel
(Marseille) sandte mir 1999 4MM und 1F einer Acalles-Art
aus dem Teno-Gebirge Tenerifes (Kanarische Inseln) zu, die ich als neue Art
neben Acalles brevitaris Wollaston 1864 von Gran Canaria und Acalles
euphorbiacus Stüben 2000 von La Palma erkannte und in meiner Monographie über
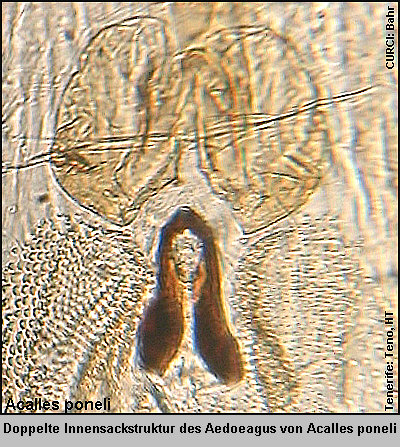
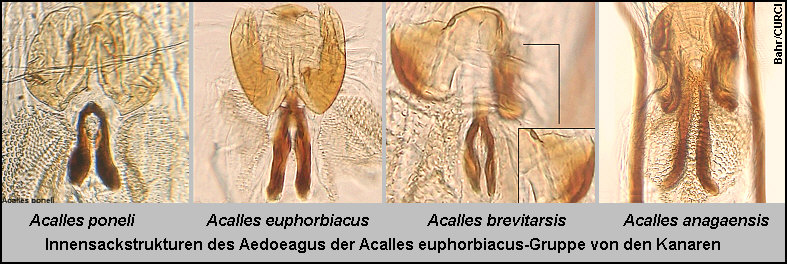
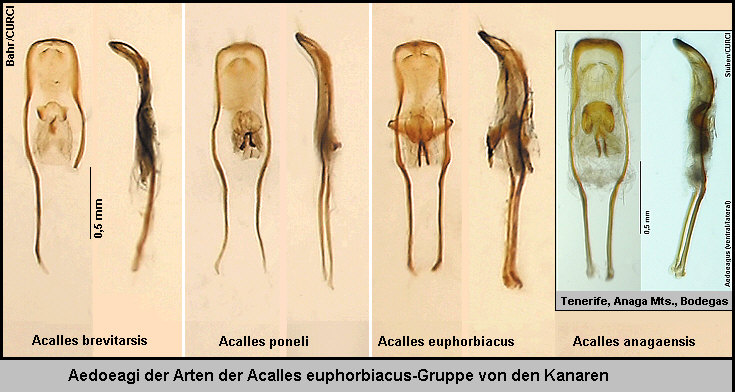
die „Cryptorhynchinae von der Kanarischen Inseln“ beschrieben habe. [Fig.
1][Fig. 2][Fig.
3][Stüben 2000e: 77-78] Die genannten drei Arten und eine
weitere Art aus dem Nordosten Tenerifes (Anaga Mts.), Acalles anagaensis
Stüben 2000, sind leicht an ihrer doppelten, sonst nur für die Arten des Laurus-Waldes
charakteristischen Innensackstruktur des Aedoeagus zu unterscheiden [Fig.
4][Fig. 5] ][Fig.
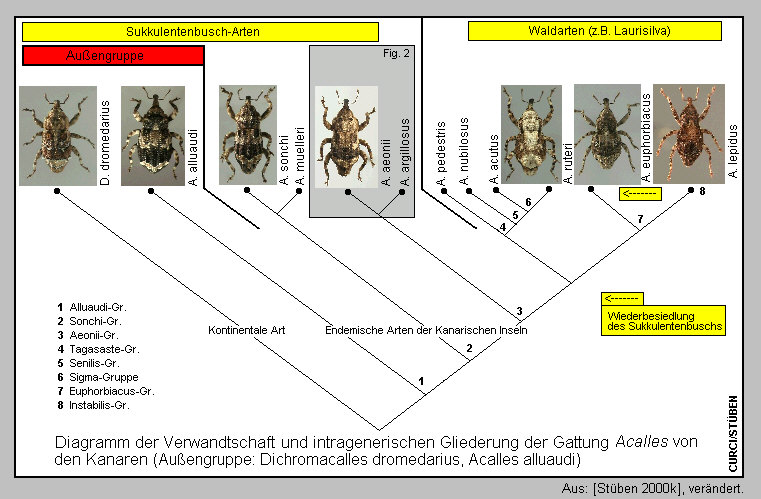
6] und gehören - meiner „Phylogenie der endemischen Taxa des Genus
Acalles von den Kanarischen Inseln“ folgend [Stüben 2000k][Germann
2004] - in die selbe Gruppe [Fig 7a]:
Die Arten der Acalles euphorbiacus-Gruppe haben sich aus den Laurus-Wäldern
kommend erneut in den oberen Sukkulentenbusch bzw. die thermophilen Buschwälder
unterhalb der feuchteren Lorbeer-Baumheide-Zone eingenischt. [Fig.
7b] Dabei galt es für drei der genannten vier Arten eine hohe Hürde
zu nehmen: die insektentoxischen Substanzen der Euphorbiaceae [Sprick & Stüben
2000] [Fig. 7c] – und für
Acalles anagaensis die ebenfalls toxisch wirkenden Inhaltstoffe einer
Boraginaceae (Echium strictum) [Germann 2004].
Erst vier
Jahre nach dem vorläufigen Abschluss meiner systematisch-taxonomischen Arbeiten
zu den Cryptorhynchinae der Kanaren besuchten Christoph Germann (CH-Bern) zum
ersten Mal und ich erneut die nördlichen Ausläufer des Teno-Gebirges im
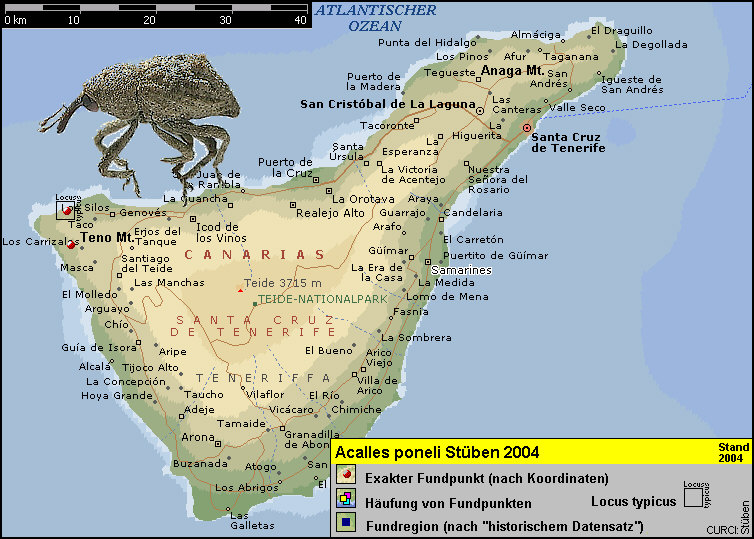
Nordwesten Tenerifes. [Fig. DaPON]
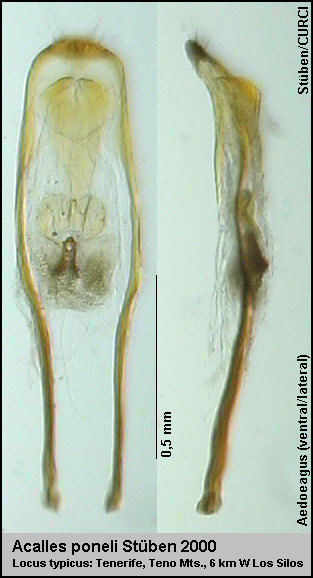
Der Locus typicus von Acalles poneli liegt 6 km westlich von Los Silos
bei Casa Blanca (N28°21'40“ W16°52'15“) mitten im Sukkulentenbusch in
einer Höhe von 120 m ü. NN. [Fig. 8]
Da dieser zweite Zyklus zu den Cryptorhynchinae der Makaronesischen Inseln in
den nächsten Jahren die Biologie, Biogeographie und Evolution der Acalles-Arten
umfassen soll, stand vom 23.12.2003 – 4.1.2004 u.a. auch die bisher weitgehend
unbekannte Biologie dieser an einer Euphorbiaceae lebenden Acalles-Art
auf dem Forschungsprogramm.
Die uns überraschenden
- wenn auch im „ökologischen Erwartungshorizont“ liegenden - Erkenntnisse
sowohl vor Ort als auch die erfolgreiche (wenn auch schwierige) Nachzucht dieser
hoch spezialisierten Acalles-Art in den Insektarien des CURCULIO
Institutes sollen hier vorgestellt werden. Darüber hinaus möchte ich erstmalig
die Eier und Larven beschreiben.
Die
Wirtspflanzenbindungen von Acalles poneli
am
Locus typicus
In der
Erstbeschreibung von Acalles poneli aus dem Jahre 2000 nenne ich, den
Angaben Philippe Ponels folgend, noch Euphorbia obtusifolia Poir. als Fraß-
und Entwicklungspflanze. Die Art lebt jedoch am Locus typicus im Nordwesten
Tenerifes an den Hängen der nördlichen Ausläufer des Teno-Gebirges auf nur
120 m Höhe an Euphorbia balsamifera Ait. [Fig.
9]. [Fig. DaPON] Dort
konnten Christoph Germann (Bern) und ich die Art zunächst in wenigen Exemplaren
des Nachts von ihrer im Absterben begriffenen Wirtspflanze im Dezember 2003 bzw.
Januar 2004 abklopfen. An gleicher Stelle wurde von uns Echinodera personata
Colonnelli 1985 in großer Anzahl aus Detritus gesiebt, eine Cryptorhynchinae,
die extrem trockene Standorte bevorzugt. [Fig.
10] Kein einziges Exemplar von Acalles
poneli fand sich an der dort ebenfalls anzutreffenden Euphorbiaceae Euphorbia
obtusifolia.
Nach
einem nächtlichen, sehr mühsamen Aufstieg des dort steil zum meeresnahen
Plateau abfallenden Teno-Gebirges traf ich in ca. 250-270 m Höhe in einer
kleinen Senke auf fünf im Absterben begriffene Exemplare von Euphorbia
atropurpurea (Brouss.) W.&B.
[Fig.
9] Hier
konnten schon nach wenigen Schlägen auf dem mitgeführten Klopfschirm etwa 1
Dutzend Tiere aufgelesen werden und das, obwohl die nächtliche Temperatur unter
10 °Celsius lag und ein leicht auffrischender Meereswind das Aufsammeln
erheblich erschwerte.
Weder in
den abgestorbenen Zweigen von Euphorbia balsamifera noch in den kräftigeren
Ästen des Tenerife-Endemiten Euphorbia atropurpurea (Dunkelpurpurrote
Wolfsmilch) fanden sich Larven oder Puppen, so dass davon ausgegangen werden
darf, dass wir es mit Exemplaren der bereits im Herbst geschlüpften letzten
Generation zu tun hatten. Da wir an vielen anderen Stellen des Teno-Gebirges
vergeblich an Euphorbia balsamifera nach Acalles poneli gesucht
haben, drängte sich schon hier - am Locus typicus - die Vermutung auf, dass es
sich bei Euphorbia atropurpurea tatsächlich um die Entwicklungspflanze
der ersten Wahl von Acalles poneli handeln könnte. Selbstverständlich
ist nicht auszuschließen, dass sich Acalles poneli auch an Euphorbia
balsamifera entwickeln kann, doch ist wahrscheinlicher, dass es sich bei den
wenigen Funden am Locus typicus eher um die letzten „Ausläufer“ einer an Euphorbia
atropurpurea lebenden Population handelt, bei der Euphorbia balsamifera weniger
als Entwicklungs-, denn als zusätzliche Fraßpflanze eine Rolle spielen dürfte.
Die
zimtbraunen Äste und der dunkelpurpurrote Blütenstand unterscheidet Euphorbia
atropurpurea leicht von den übrigen strauchförmigen Euphorbien. [Fig.
11] Die Pflanze ist im Teno-Gebirge
keineswegs häufig. Sie präferiert felsige, in der Regel küstenferne Standorte
im oberen Sukkulentenbusch und steigt höher hinauf als ihre verwandten Arten. [Fig.
12]
Acalles
poneli und die
Entwicklungspflanze
Euphorbia
atropurpurea
Am
2.1.2004 entdeckten wir im äußersten Westen des Teno, westlich der kleinen
Ortschaft Los Carrizales (Bco. del Carrizal) Acalles poneli auf den im
Absterben begriffenen Zweigen von Euphorbia atropurpurea. [Fig.
13][Fig.
DaPON] Die von dieser Cryptorhynchinae
befallenen Pflanzen standen am Fuße einer Felswand auf einem sehr feuchten
Quellhang in einer Höhe von 370 m ü NN. [Fig.
14]. Uns fiel gleich auf, dass unter den
zahlreichen Exemplaren ein hoher Prozentsatz abgestorben bzw. im Absterben
begriffen war. Offensichtlich handelte es sich um eine Population, die in den
Wintermonaten regelmäßig einem Feuchtigkeitsstress ausgesetzt ist, sich aber
in den wärmeren und vor allem trockeneren Monaten des Jahres wieder erholt. [Fig.
15]
Die Acalles
poneli-Exemplare von den zimtbraunen Ästen dieser strauchförmigen
Euphorbiaceae sind viel kontrastreicher gezeichnet und deutlich größer als die
Exemplare, die von uns am Locus typicus (vor allem an Euphorbia balsamifera)
in den Tagen zuvor westlich von Los Silos nachgewiesen wurden. Sie gehören
aber zweifelsfrei zur selben Art! [Fig.
2]
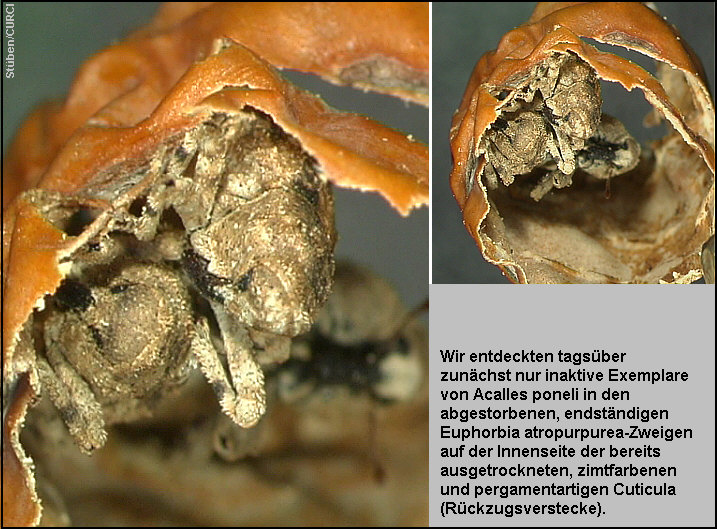
Ich
entdeckte zunächst in den späten Nachmittagsstunden ein Exemplar in einem
abgestorbenen, endständigen Euphorbia atropurpurea-Zweig zwischen dem
noch harten, röhrenfömigen Splintholz und der bereits ausgetrockneten,
zimtfarbenen pergamentartigen Rinde. [Fig.
16] Weitere Exemplare wurden in ihren
offensichtlich nur tagsüber aufgesuchten Verstecken unterhalb der schopfig an
den Zweigenden angeordneten, bisweilen purpurrot überlaufenen Blätter, den
hellgrauen Abwurfstellen der Blätter und den Zweiggabelungen entdeckt. [Fig.
17] Erst in den Nachtstunden fanden wir
dann in großer Anzahl Acalles poneli auf den im Absterben begriffenen
Zweigen herumlaufen. [Fig. 18]
Trotz der tiefen Temperaturen (weit unter 10 °Celsius) kopulierten die Tiere
und fraßen von der abgestorbenen pergamentartigen Rinde. Auch auf dem
Klopfschirm verharrten die Tiere nicht lange in der sonst üblichen Katalepsie,
sondern zeigten ein hochaktives Verhalten. Auch hier wurden von uns keine Larven
oder Puppen gefunden.
Von
diesem Standort (E.: Tenerife, Teno Mts, 8 km NW Santiago del Teide, W Los
Carrizales, N28°19'14"W16°52'03", 370m) wurden etwa zwei Dutzend
Exemplare von Acalles poneli und frische Zweige von Euphorbia
atropurpurea zur Nachzucht mit ins CURCULIO-Institut nach Mönchengladbach
(Germany) gebracht.
Die
erfolgreiche Nachzucht
von
Acalles poneli an Euphorbia atropurpurea
Die
Zucht von Acalles poneli an Euphorbia atropurpurea sollte sich als
schwierig erweisen. Was die Nachzucht an Euphorbiaceae betrifft, so lagen mir
bisher nur eigene Erkenntnisse von erfolgreichen Zuchten an Euphorbia
nicaeensis (Kyklioacalles euphorbiophilus Stüben 2003, Marokko) und Euphorbia
stygiana (Callacalles droueti; Azoren) vor. [Stüben 2003d][Stüben
2004a] Allerdings handelte es sich dabei um Pflanzen, die entweder aufgrund
ihrer schlanken, langen Zweige - wie im Fall von Euphorbia nicaeensis -
rasch austrocknen oder – wie im Fall der baumförmigen Euphorbiaceae Euphorbia
stygiana – nach Wasserentzug sehr schnell aushärten. Beides trifft, wie
sich zeigen sollte, für die 2-2,5 cm starken, sehr „vitalen“ Zweige von Euphorbia
atropurpurea nicht zu: Noch nach fünf Monaten
trat nach Abbruch der letzten endständigen Blätter klebriger Milchsaft aus.
Der Milchsaft der Euphorbiaceae enthält u.a. reizende Harze, Diterpene, tetra-
und pentacyclische Triterpene, ichthyozide Ester, z.B. das cocarcinogene
Phorbol, Estersäuren wie Tiglin- oder Phenylessigsäure und ungesättigte Fettsäuren
([Hegnauer 1966][Hegnauer 1989]) und weist somit eine große Fülle biologisch
aktiver und sicher auch mehr oder weniger stark insektentoxischer Substanzen
auf; also denkbar schlechteste Voraussetzungen für eine erfolgreiche larvale
Entwicklung, die nur einsetzen kann, wenn sich die insektentoxischen Substanzen
nach dem Absterben der Pflanze weitgehend abgebaut haben. [Sprick & Stüben
2000] Außerdem erhöht ein schleppender Austrocknungsprozess das Risiko einer
vorzeitigen Verpilzung.
Zuchtansatz
(7.1.2004)
Am 7.1.2004
wurden von mir 6 abgeschnittene frische Zweige von Euphorbia atropurpurea mit
den schopfigen Blättern an den Zweigenden senkrecht in ein mit einer 2 cm hohen
Sandauflage gefülltes Glas-Insektarium eingebracht. [Fig.
19] Die etwa 2-2,5 cm dicken Zweige waren zwischen 15 cm und 25 cm
lang. Dazu wurden 8FF und 8MM von Acalles poneli vom oben
genannten Fundort im Teno-Gebirge gesetzt. In den ersten beiden Wochen wurden in
den dem Sandboden aufsitzenden, feuchteren Endsegmenten der sehr langsam
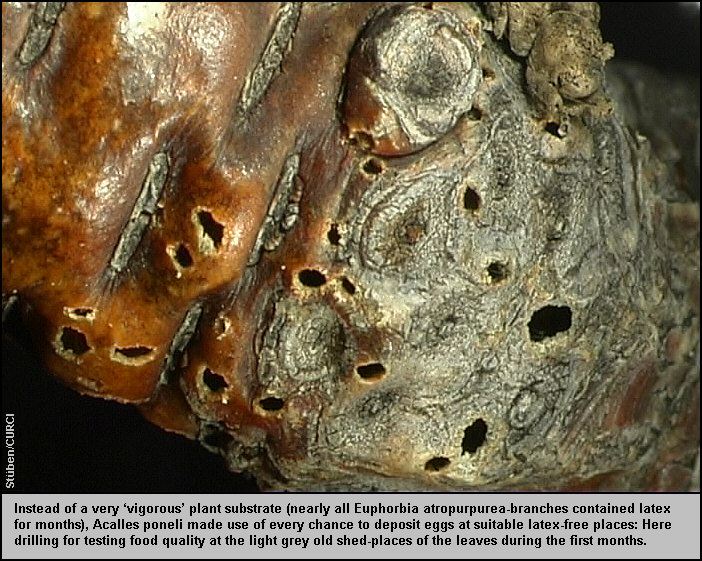
austrocknenden Zweige winzige Löcher in die pergamentartige, zimtbraune Rinde
gefressen. (Dabei könnte es sich um erste Testlöcher für die Eiablage
gehandelt haben). [Fig. 20]
Nach 22.00 Uhr laufen die sich tagsüber unter den abgestorbenen, schopfartig
herunterhängenden Blättern oder in kleine Hohlräume zurückziehenden Tiere [Fig.
21] auf den Hauptzweigen herum und kopulieren. Die Raumtemperatur
betrug tagsüber zwischen 22 ° und 24 °Celsius; nachts wurden Temperaturen
zwischen 18 ° und 20 °Celsius gemessen.
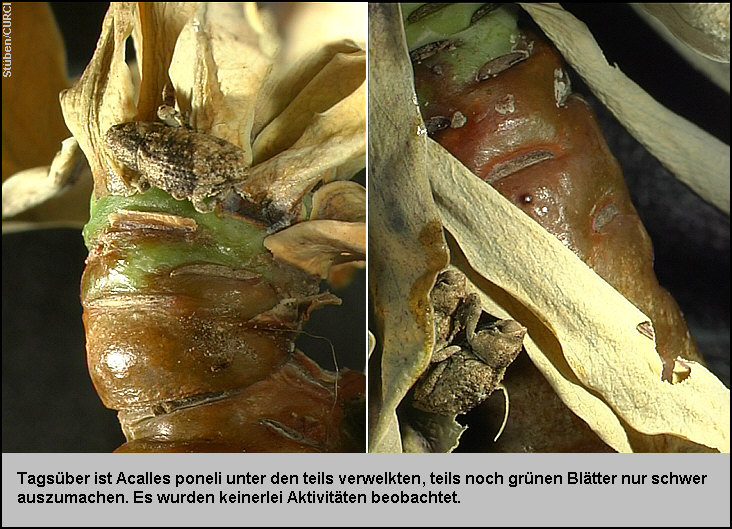
Tagsüber sind
die Tiere unter den teils verwelkten teils noch grünen Blättern nur schwer
auszumachen und es werden keinerlei Aktivitäten beobachtet. [Fig.
22] In den Nachtstunden werden die welken Blätter bevorzugt
aufgesucht [Fig. 23], und
die Tiere zeigen auf den senkrecht platzierten Zweigen ein hoch aktives
Laufverhalten. [Fig. 24]
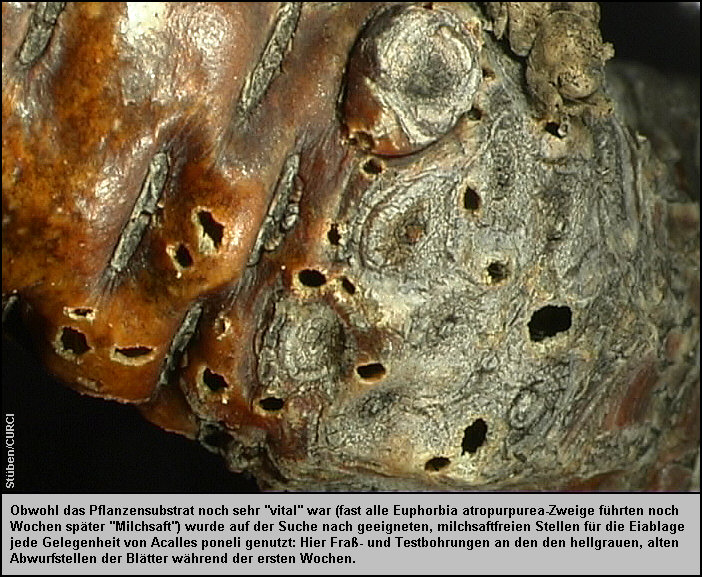
Lochfraßspuren fanden sich auch an den Zweiggabelungen bzw. an den hellgrauen
Abwurfstellen der Blätter aus den letzten Entwicklungsjahren der Pflanze. [Fig.
20] Vier Wochen nach dem Eintrag wurden an einer solchen Abwurfstelle
zwischen der pergamentartigen Rinde und dem harten Schaftholz zwei Eier entdeckt
(Länge:1,13 mm, Breite: 0,88 mm). Die Eier steckten zur Hälfte in dieser etwa
2 mm dicken, zum Teil schon stark verpilzten (vermatschten) Zwischenschicht. [Fig.
25][Fig. 26]
Larven wurden noch nicht entdeckt.
Offensichtlich
aber waren die Zweige immer noch zu frisch, denn nach Abnahme der grünen Blätter
oder nach dem Anschneiden der Zweige trat immer noch weißer Milchsaft aus. Bis
zu diesem Zeitpunkt wurden die Euphorbia atropurpurea-Zweige alle 2-3
Tage besprüht, da mir aus vorangegangenen Zuchten an Euphorbiaceae bekannt war,
dass „genügend Feuchtigkeit“ in den ersten beiden Larvalstadien der
entscheidende Parameter für eine erfolgreiche Entwicklung ist. Andererseits
vertragen Cryptorhynchinae-Larven im letzten Entwicklungsstadium, also nach
ihrem „Abtauchen“ ins härtere Splintholz, praktisch keinen
Feuchtigkeitseintrag mehr. Insofern waren die ersten Wochen eine Gratwanderung
zwischen einer notwendigen Austrocknung und damit des Abbaues der Milchsaft führenden
Schichten bzw. insektentoxischen Substanzen und dem Erhalt eines Minimums
an Feuchtigkeit in den ersten Larvalstadien.
2.
Visite (5.3.2004)
Zu diesem
Zeitpunkt wurden von mir 2 Larven im 1. Larvenstadium (Maße: 2,2 mm x 0,7mm)
und eine weitere Larve im 2. Larvenstadium im (meines Erachtens immer noch viel
zu feuchten) matschigen Pflanzensubstrat unterhalb der pergamentartigen Rinde,
aber oberhalb des verholzten Schafts entdeckt. [Fig.
27][Fig. 32]
Die eingetragenen Imagines schienen - da ich keine toten Exemplare ausmachen
konnte - noch alle vollzählig zu sein und mit den Bedingungen im Insektarium
hervorragend auszukommen. Nach den Besprühungen, die von mir vor allem in den
Abendstunden vorgenommen wurden, zeigten die Imagines auf den Zweigen immer noch
ein hoch aktives Lauf-, Fraß- und Kopulationsverhalten. Dennoch stellte ich ab
diesem Zeitpunkt die wöchentlichen Besprühungen völlig ein, um den Larven
eine Entwicklungsmöglichkeit zu geben!
3.
Visite: (4.4.2004)
Von den 16
Ausgangstieren konnten 13 Exemplare und 1 totes Exemplar ohne großen Aufwand
(also ohne Zerstörung des Pflanzenmaterials) wieder entdeckt werden. Alle
lebenden Tiere wurden ins Insektarium zurückgegeben. Eine Larve konnte im 3.
Larvenstadium (Maße: ca 5,5 mm x 1,5 mm) dem weitgehend zerraspelten Mark im
Inneren eines ausgetrockneten Zweiges entnommen werden. [Fig.
28][Fig. 33]
Gesamtsituation
zu diesem Zeitpunkt: Obwohl die Zucht jetzt bereits drei Monate andauerte,
konnten weder Puppen noch zahlreiche Larven entdeckt werden. Dies legt den
Schluss nahe, dass die Euphorbia atropurpurea-Zweige, die z.T. noch immer
sehr „vital“ sind, also Milchsaft führen, den Imagines in diesem Zustand
praktisch kaum eine Chance zur Eiablage bieten. So konnten nur an wenigen,
milchsaftfreien Stellen Eiablagen stattfinden und die Entwicklung der Larven
einsetzen. Aus diesem Grund habe ich die 13 noch lebenden Elterntiere wieder
eingebracht. Für die „verspätete“ Eiablage spricht auch, dass in den
ersten 6 Wochen unter der pergamentartigen Cuticula z.T. Pilzbefall einsetzte,
die Gesamtsituation also zu feucht war. Diese hatte sich durch das Wegfallen der
Besprühungen und der wesentlich höheren Raumtemperatur (tagsüber in der
letzten Woche 25 °-27 °Celsius) merklich verbessert.
4.
Visite (9.5.2004)
In den jetzt
hohlen, ausgetrockneten Zweigen wurden die Überreste des Raspelspäne-Fraßes
der Larven (und ihrer Ausscheidungen) entdeckt. Dort konnten insgesamt 3 Larven
im letzten Larvenstadium entdeckt und photographiert werden [Fig.
28] Die Larven leben jetzt im Inneren der röhrenartigen Schafte,
nachdem sie den schmalen Zwischenraum unterhalb der pergamentartigen Rinde
verlassen und sich durch das harte Schaftholz gefressen haben (doppelter
Schutz). Es wurde auch ein Schlupfloch entdeckt und photographiert. [Fig.
29] Ebenso wurde eine Puppe (noch ohne Ausfärbung der Augen) in
ihrer Puppenwiege entdeckt und zur weiteren Entwicklung separiert! [Fig.
29]
Am 16.5.2004
wurden Fotos von den Überresten der Pflanzen gemacht, in denen sich auch
weiterhin Larven entwickelten. [Fig.
30]
Die separierte
Puppe hat sich zur fertigen Imago zwischen dem 20. und 22.5.2004
entwickelt. Das Puppenstadium wird infolgedessen etwa 2 Wochen umfasst haben (für
eine Acalles-Art ungewöhnlich lang!). Am 24.5.2004 war das Exemplar in
der Puppenwiege völlig ausgefärbt, aber noch nicht ganz ausgehärtet. Durch
das vorsichtige Erweitern des beim Aufbrechen der Puppenwiege am 9.5.2004
entstandenen kleinen „Schlupfloches“ wurde die Imago zum Verlassen der
Puppenwiege bewogen. [Fig. 31]
Wieder konnte die Imago in der „steinharten“ Hülle des Euphorbia
atropurpurea-Zweiges nicht dabei beobachtet werden, dass sie zu diesem
Zeitpunkt in der Lage gewesen wäre - oder Anstalten machte - sich selbst zu
„befreien“ (vgl. [Stüben 2003d][Stüben 2004a]) So
darf angenommen werden, dass auch dieses Exemplar noch Wochen in dem sicheren
Schaft verharrt hätte, wenn nicht eine vorzeitige „mechanische Befreiung“
eingeleitet worden wäre. Erst nach völliger Aushärtung des Rüssels und nach
Aufweichung des Zweiges durch Wind und Wetter wird wohl der eigentliche Schlüpfvorgang
auf Tenerife (im Spätherbst ?) einsetzen.
Zuchtende:
1.6.2004)
In den bereits
am 16.5.2004 photographierten Überresten der an vielen Stellen aufgebrochenen
Zweige [Fig. 30] konnten
beim „Ausräumen“ des Insektariums 12 Elterntiere, 1 totes Exemplar, 1
frisch geschlüpfte, noch weiche Imago und insgesamt 4 weitere Larven entdeckt
bzw. wieder entdeckt werden.
1.
Da auch diese Zucht nach einer sehr aufwendigen „try and
error“-Methode stattfand (die jedoch - im Gegensatz zu den üblichen
Serien-Zuchten „in vitro“, z.B. in Petrischalen
[Stüben 2004a] - in geräumigen Insektarien fast immer auch den Erfolg einer
Cryptorhynchinae-Zucht garantiert), also jederzeit eingreifende
Korrekturen in die Zusammensetzung der den Metabolismus bestimmenden
„Umwelt“-Parameter vorgenommen werden, kann über den genauen Zeitraum der
postembryonalen Entwicklung (Metamorphose) keine definitve Aussage gemacht
werden; zumal Cryptorhynchinae zu den endophytischen Holz- und Wurzelbohrern
gehören und daher unter gewissen Verlusten an Pflanzensubstrat nur regelmäßige
„Visiten“ (etwa in 4wöchigen Abständen) in Frage kommen.
Geht man jedoch davon aus, dass sich während der ersten 6 Wochen des
Experimentierens in dem noch viel zu feuchten und Milchsaft führenden, also
in dem noch immer insektentoxischen Substrat, keine Larven entwickelt haben dürften
und zieht den Schlüpfzeitpunkt der ersten Imago am 22.5.2004 heran, dann
darf man wohl von einem ca. 3 Monate
umfassenden Entwicklungszeitraum -
von der Eiablage bis zur Imago - ausgehen. Eine Annahme, die von allen
bisherigen Erfahrungen bei der Zucht von Cryptorhynchinae, insbesondere von
den Arten der Gattungen Calacalles, Acalles und Kyklioacalles,
bestätigt wird! [Stüben 2003d][Stüben 2004a]
2.
Spannend bleibt natürlich die Frage nach dem jahreszeitlichen
Entwicklungszeitraum in natura. Dazu sind die Beobachtungen und die Fundumstände
von Acalles poneli und der Entwicklungspflanze Euphorbia
atropurpurea im äußersten Westen des Teno-Gebirges auf Tenerife
heranzuziehen. Die von dieser Cryptorhynchinae
befallenen Pflanzen standen am Fuße einer Felswand auf einem im Winter
2003/2004 sehr feuchten Quellhang des Barranco del Carrizal. Zu diesem
Zeitpunkt führte der Barranco del Carrizal noch Wasser, das in den heißeren
Sommermonaten völlig versiegt. Dann erreichen auch die von Norden kommenden
Passatwolken nur noch an wenigen Tagen den westlichen Zipfel des Teno-Massivs.
Alle diese Faktoren zusammen erklären den hohen Prozentsatz abgestorbener Euphorbia
atropurpurea-Sträucher auf dem zumindest in den Wintermonaten sehr
feuchten Quellhang: Der harte Wechsel
von extrem feuchten zu extrem trockenen Habitatbedingungen schafft eine
ausgesprochen anfällige „Stresspflanzen“-Population, also ein ideales
Areal für endophytische Lebensformen.
Da wir weder Larven noch Puppen im Winter 2003/2004 vorfanden und mit einer
Eiablage zu diesem Zeitpunkt schon deshalb nicht zu rechnen ist, weil die
(nicht bereits im letzten Jahr völlig abgestorbenen) stark Milchsaft führenden
Euphorbia atropurpurea-Sträucher angesichts des ausgiebigen
Wasserangebots in den Wintermonaten sehr „vital“ sind (sich erholen!),
muss bei Acalles poneli von einem Entwicklungszeitraum in den
trockeneren Sommermonaten (mit einem Maximum im Juni - August) ausgegangen
werden. Mit dem völligen Trockenfallen des felsigen Hanges werden zahlreiche
Pflanzen absterben, ein Prozess, der wie die Zucht an frischen Euphorbia
atropurpurea-Zweigen gezeigt hat, sehr frühzeitig im Jahr (April?)
einsetzen muss, denn noch lange behalten selbst abgetrennte Zweige unter der
zimtbraunen Rinde ihren insektentoxischen Milchsaft.
3. Die Existenz der Wirtspflanze Euphorbia atropurpurea ist zwar eine notwendige , nicht aber schon hinreichende Voraussetzung für die Eiablage und für die daran anschließende Entwicklung zur fertigen Imago: Denn nicht so sehr das Vorhandensein der Wirtspflanze Euphorbia atropurpurea an sich ist entscheidend, sondern der rasche Wechsel von extrem feuchten zu extrem trockenen Habitatstrukturen („Wechselhabitate“ / „switch-habitats“) ist für den Entwicklungszyklus von Acalles poneli ausschlaggebend! Dies deckt sich mit zwei Beobachtungen: A. Der oben genannte Quellhang an einer steilen Felswand geht nach Osten in eine noch weitaus größere, das ganze Jahr über relativ trocken stehende Euphorbia atropurpurea-Population über. Hier fanden sich nur noch sehr wenige Exemplare von Acalles poneli (Verhältnis: ca. 10:1). B. Auch die oben beschriebene Zucht hat es wieder unter Beweis gestellt: „Genügend Feuchtigkeit“ ist in den ersten beiden Larvalstadien der entscheidende Parameter für eine erfolgreiche Entwicklung von Acalles – hier: zwischen der pergamentartigen Rinde und dem harten, splintholzartigen Schaft. Erst die Larven im letzten, dritten Stadium verlassen den „matschigen“, dunkelbraunen bis schwarzen „Zwischenraum“ und fressen sich durch den harten Schaft in das zu diesem Zeitpunkt schon weitgehend ausgetrocknete Schaftmark [Fig. 27], wo sie sich schließlich in der aus trockenen Raspelspänen „gestauchten“ (bzw. „gestampften“) Wiege verpuppen. [Fig. 29] In dieser Phase sind die Larven und Puppen auf extrem trockene Habitatbedingungen angewiesen!
4. Grundsätzlich - und auch dies zeigte die Nachzucht von Acalles poneli - sind unsere flugunfähigen Cryptorhynchinae bei der Eiablage nicht an feste Jahreszeiten gebunden. Auf den Kanaren sind die jahreszeitlichen Temperaturschwankungen ohnehin sehr ausgeglichen und im nordöstlichen Las Mercedes-Wald des Anaga-Gebirges gibt es keinen Monat ohne Niederschläge (mit sehr hohen Werten von über 150 mm von Dezember bis Februar). Wo immer sich die Möglichkeit bietet, sei es z.B. durch ein stressbedingtes Absterben der Pflanzen (wie im geschilderten Fall) oder durch mechanische Einwirkungen (wie z.B. bei partiellem Wind- oder Geröllbruch), werden die Tiere sofort(!) nach Abriss (Zerfall) der toxischen Pflanzeninhaltstoffe mit der Eiablage beginnen. Dass sie als Entwicklungseinstieg allerdings eine Phase präferieren, die zunächst noch in einer Feuchtigkeitsperiode liegt, dann aber spätestens mit dem Beginn des dritten larvalen Stadiums in eine möglichst extreme Trockenperiode umschlägt, muss man unterstellen. Es zum Beginn einer Feuchtigkeitsperiode zu „wagen“, also Eier abzulegen, verbietet allein schon die Gefahr von Staunässe unter der pergamentartigen, Evaporation weitgehend unterbindenden Cuticula vieler Euphorbiaceae, die zu einer vorzeitigen Verpilzung der Larven führen könnte. Umgekehrt scheidet eine Eiablage auf dem Höhepunkt einer Trockenphase aus den gleichen Gründen aus. Aber noch niemals wurden Acalles-Arten dabei beobachtet, dass sie an Trocken- oder Totholz eine Eiablage versucht hätten! (Auch wenn dies in völliger Verkennung der unabdingbaren, komplexen biotischen und abiotischen Voraussetzungen einer erfolgreichen Metamorphose bei Cryptorhynchinae immer wieder behauptet wird. [Kippenberg 1983:162] [Koch 1992: 301])
Beschreibung der Eier und Larven von Acalle poneli
Ei:
Zwei Eier wurden von mir unmittelbar nach der Eiablage unterhalb der
zimtfarbenen Cuticula einem Euphorbia atropurpurea-Zweig entnommen und
untersucht. [Fig. 25][Fig.
26] Das kurzovale Ei ist weiß, glatt, 1,13 mm lang und 0,88 mm
breit.
Larve
(letztes Larvenstadium): [Fig.
33][Fig. 32]
Habitus:
Größe: Länge 5,5 – 6,0 mm; Breite ca. 1,5 – 2,0 mm. Körper weiß, weich
(verformbar) ohne auffallende Sklerotisierungen, sowohl im lebenden als auch im
(in Fixierlösung) abgetöteten Zustand stets mehr oder weniger stark ventralwärts
bogig gekrümmt, beinlos. Segmentierung deutlich. Kopfkapsel hellbraun bis
dunkelbraun gefärbt, stärker chitinisiert. Gesamte Körperoberfläche glänzend
und mehr oder weniger kahl erscheinend; nur auf der Unterseite mit Borsten und
Sensillen spärlich besetzt.
Kopf:
Die braune Kopfkapsel ist je nach dem Grad ihrer Chitinisierung unterschiedlich
stark angedunkelt. Während die am stärksten beanspruchten Spitzen der
Mandibeln oft schwarz erscheinen, ist das Labium oft nur noch teilweise
chitinisiert und entsprechend aufgehellt. Die gesamte Oberfläche des Kopfes und
der Mundwerkzeuge ist unbehaart und nur mit einzelnen, weit voneinander entfernt
stehenden bzw. zu Gruppen zusammentretenden Borsten und Sensillen ausgestattet.
Seiten des Kopfes glatt und gleichmäßig gerundet, zum Vorder- und Hinterende
hin schwach konvergierend; Frons abgeflacht; Mandibeln von oben sichtbar. Am
seitlichen Vorderrand (der Frons) liegen kleine und undeutliche, eingliedrige
Antennen, diese an der Spitze mit feinen Sensillen. Die rudimentierten Augen der
Larve grenzen direkt an den Außenrand der Antennen an.
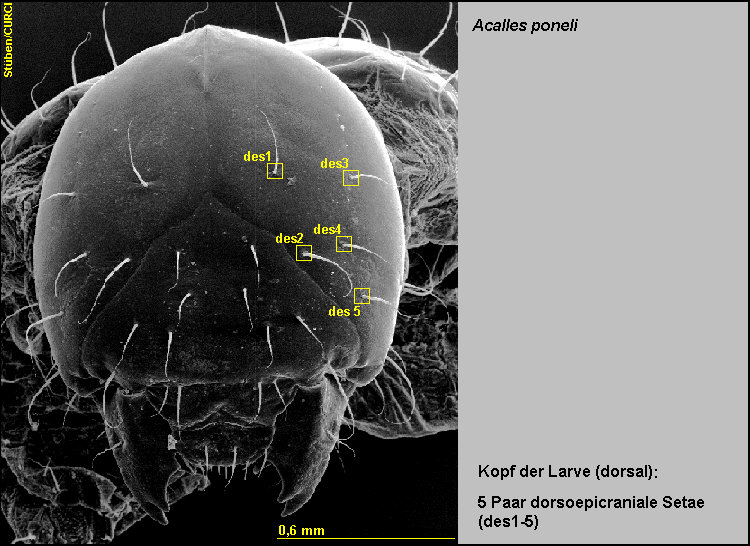
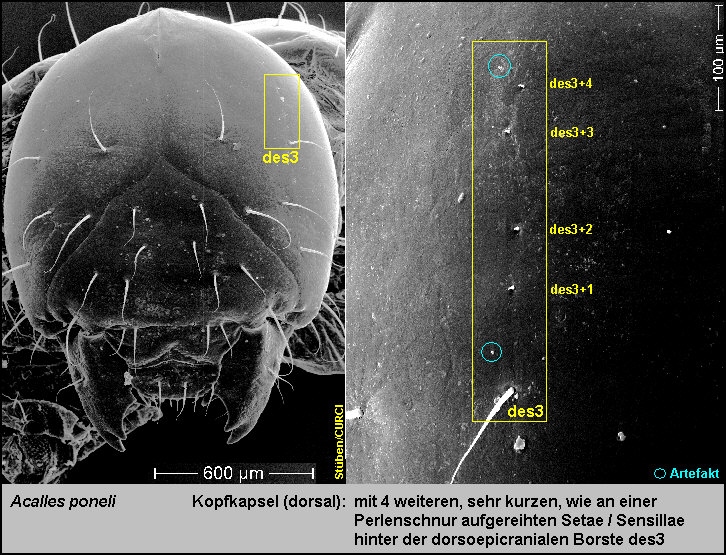
Beborstung
des Kopfes
Die Kopfkapsel trägt:
-
5
Paar dorsoepicraniale Setae (des1-5) [Fig.
LPon1]
mit 4 weiteren, sehr kurzen Sensillae [Fig.
Lpon11],
-
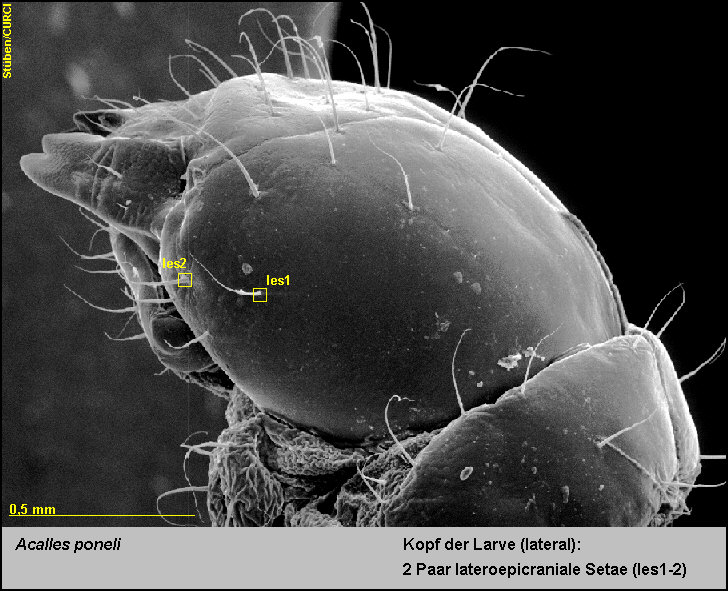
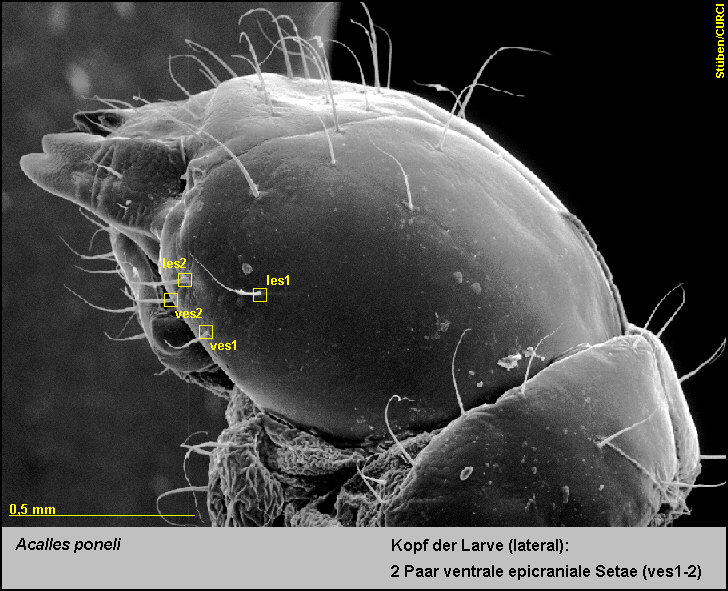
2 Paar
lateroepicraniale Setae (les1-2) [Fig.
LPon2]
und
-
2 Paar ventrale epicraniale Setae (ves1-2) [Fig.
LPon3].
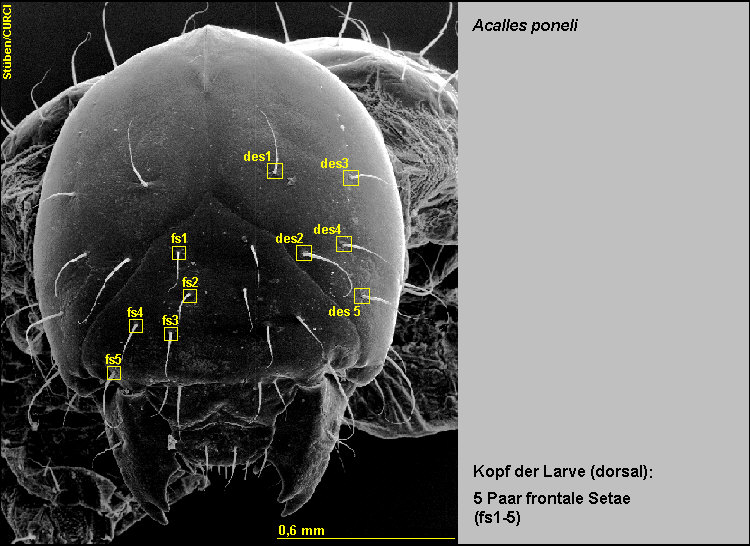
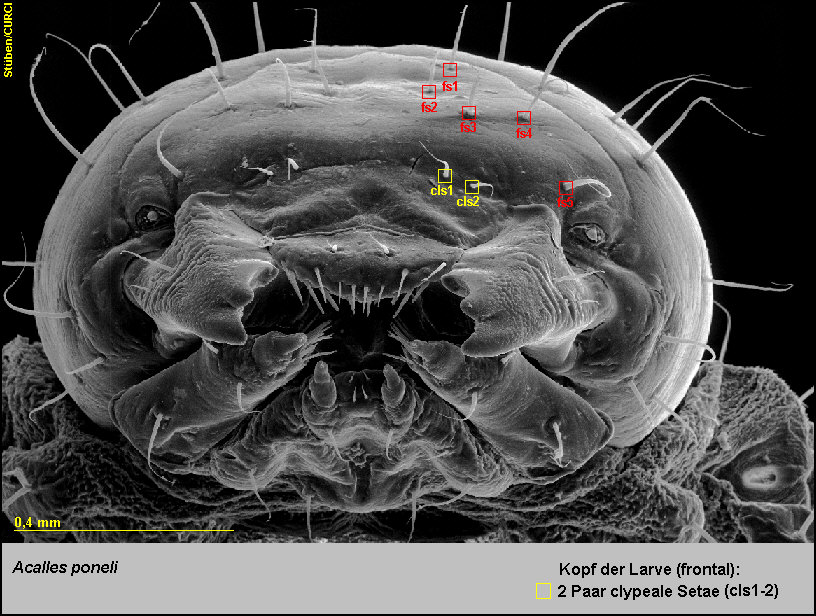
-
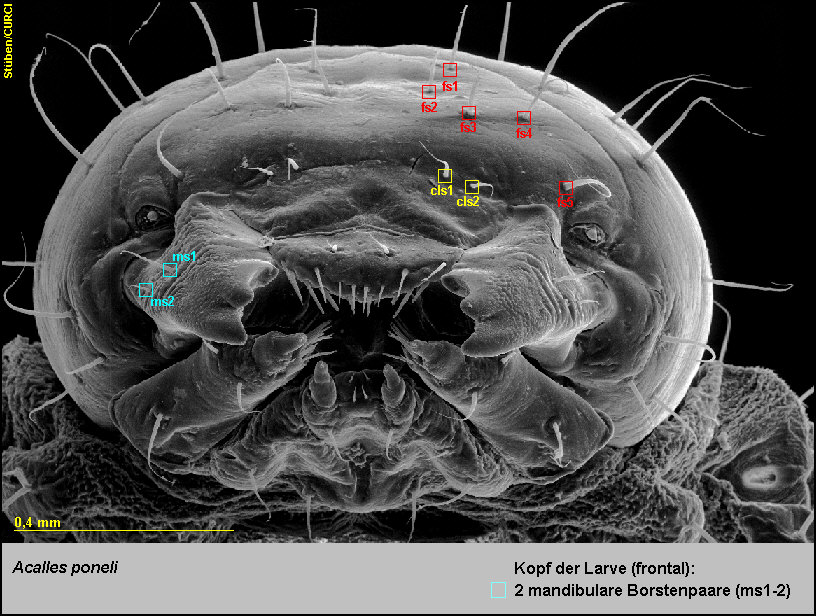
Die Stirn ist mit 5 Paaren frontaler Setae (fs1-5) ausgestattet [Fig.
LPon4].
-
Der Clypeus trägt am Hinterrand 2 Paare clypealer Setae (cls1-2) [Fig.
LPon5].
-
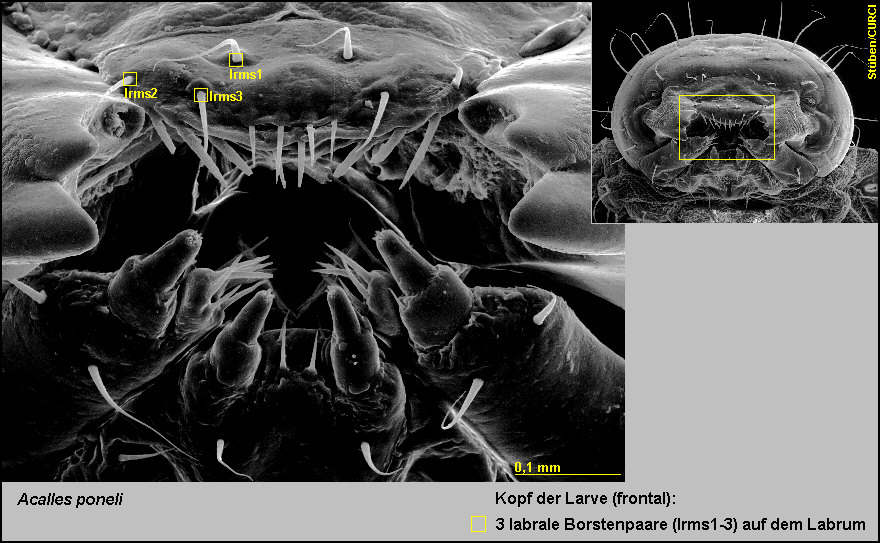
Das Labrum ist auf seiner Oberfläche mit 3 labralen Borstenpaaren
(lrms1-3) versehen [Fig.
LPon6].
-
Die Außenflächen der Mandibeln tragen je 2 mandibulare Setae (ms1-2) [Fig.
LPon7].
Der
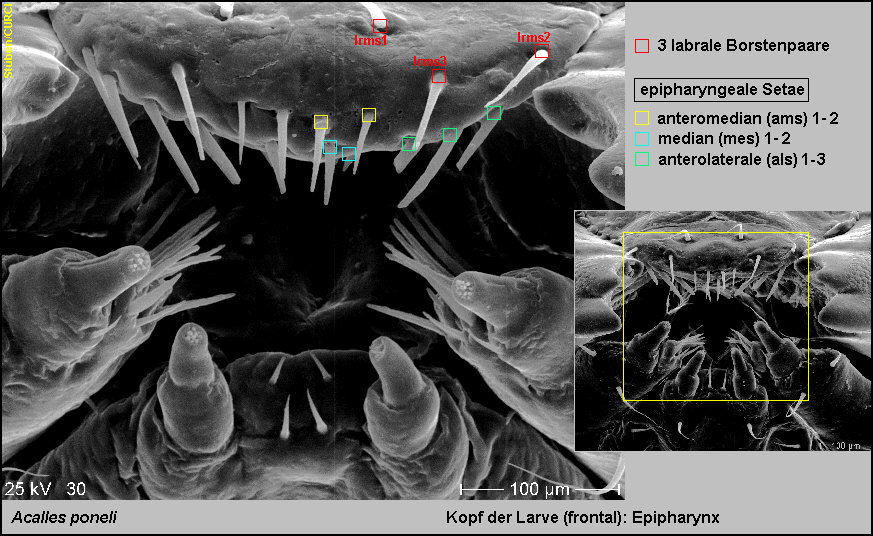
Epipharynx trägt:
-
2 Paar anteromediane Setae (ams1-2),
-
2 Paar mediane Setae (mes1-2) und
-
3 Paar anterilaterale Setae (als1-3) [Fig.
Lpon8].
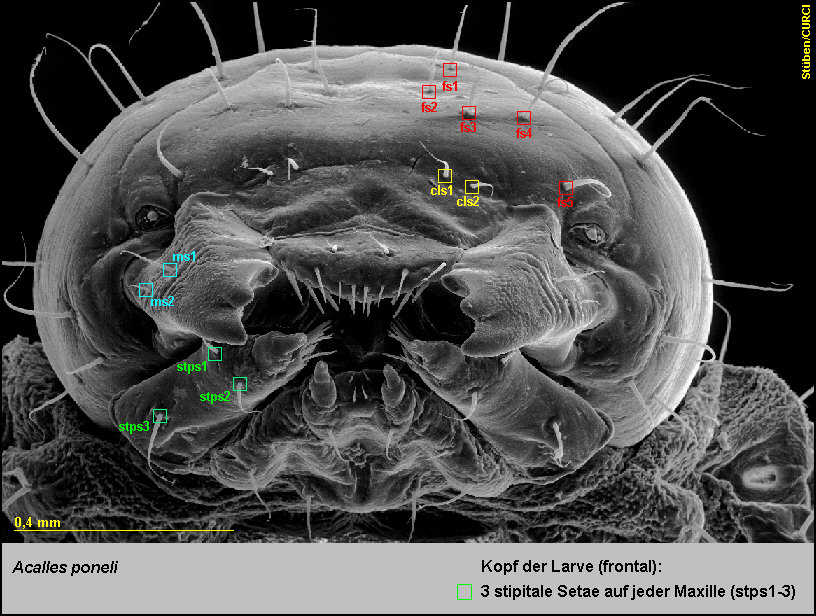
Jede
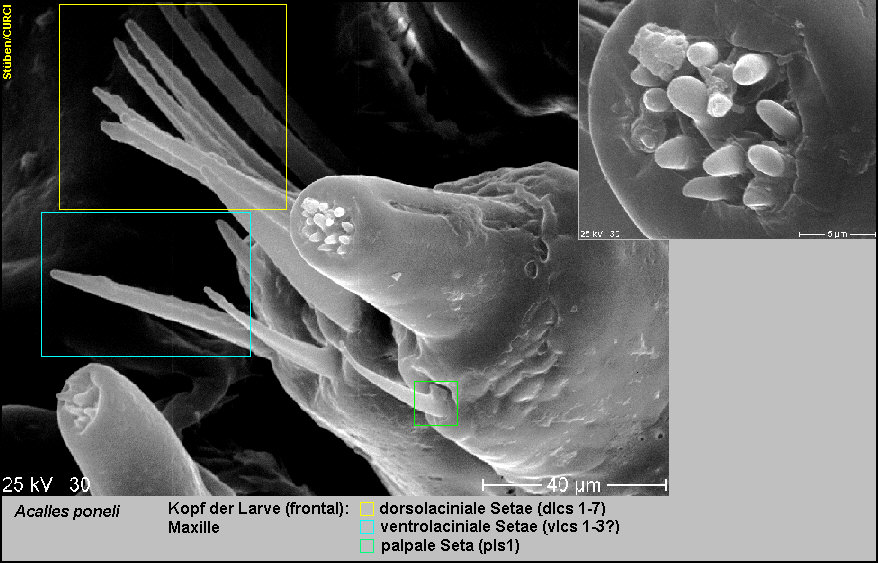
Maxille trägt:
-
3 stipitale Setae (stps1-3) [Fig.
LPon9],
-
7 dorsolaciniale Setae (dlcs1-7),
-
3(?) ventrolaciniale Setae (vlcs1-3?) und
-
1
palpale Seta (pls) [Fig.
LPon9a].
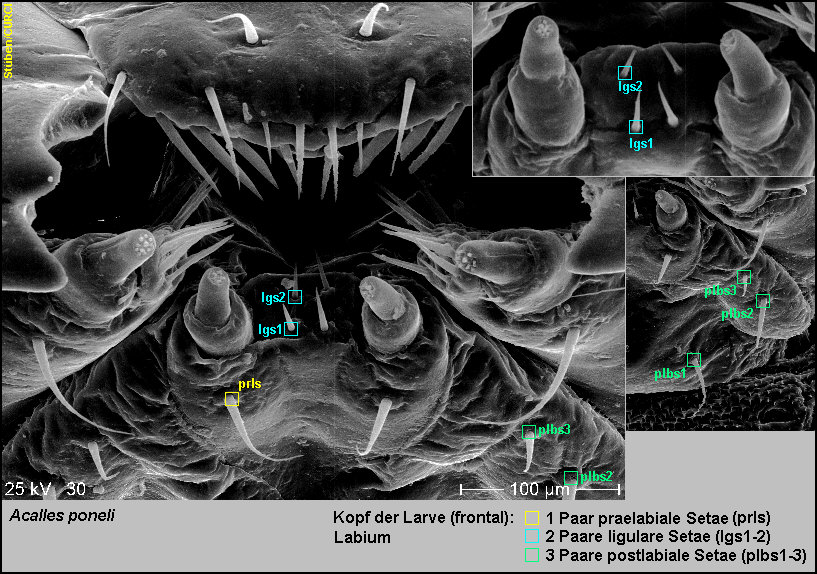
Das
Labium trägt:
-
1 Paar praelabilae Setae (prls),
-
2 Paar ligulare Setae (lgs1-2) und
-
3
Paar postlabiale Setae (plbs1-3) [Fig.
LPon10].
Diskussion
und Differentialdiagnose:
Die Beschreibung der Larven von Acalles poneli Stüben 2000 orientiert sich an den Erstbeschreibungen der kanarischen Acalles- und Dichromacalles-Larven von Ch. Bayer [Bayer & Stüben 2000] und meinen Erstbeschreibungen der Larven von Kyklioacalles bupleuri (Tunesien) und Kyklioacalles euphorbiophilus (Marokko) [Stüben 2004b, in print] sowie der Larven-Beschreibung zweier weiterer Calacalles-Arten von den Azoren [Stüben 2004d, in print].
Grundsätzlich gilt (nach dem derzeitigen Stand der Forschung) für alle Acalles-, Dichromacalles- und Calacalles-Larven:
1. Der direkte, interartliche Vergleich der Larven zeigt keine signifikanten Unterschiede sowohl was die Anzahl als auch die Lage der Setae und Sensillae betrifft.
2. Schon intraspezifische Vergleichsserien von Larven der selben Art in einem vergleichbaren Entwicklungsstadium (unmittelbar vor der Verpuppung) lassen eine rein „metrische“ Vorgehensweise mehr als fragwürdig erscheinen. Die ausschließlich endophytische Lebensweise der Cryptorhynchinae-Larven in mulmigem oder faserigem Rinden- oder Splintholz führt regelmäßig zu Deformationen, zu Stauchungen und zu die Entwicklung störenden kleinen „Unfällen“ (Alterung). So verlieren die Larven Borsten, die Mandibeln zeigen im Rasterelektronenmikroskop Risse, die Kopfkapsel weist Deformationen auf und gelegentlich tauchen sogar vereinzelte, nicht-paarig angelegte Setae oder Sensillae auf. Daher macht es wenig Sinn, die Lage der Borsten zueinander „metrisch“ zu fixieren, um artspezifische Ergebnisse für den interartlichen Vergleich zu erhalten. (Anmerkung: Die optische Ausrichtung der abgeflachten Kopfkapsel zu Vergleichszwecken am Rasterelektronen-Mikroskop ist sehr schwierig, da schon wenige Hundertstel Millimeter teils erhebliche Positonsverschiebungen der Borsten suggerieren.)
3. Da die Anzahl und die relative Lage der Borsten innerhalb einer Cryptorhynchinae-Gattung für die unterschiedlichen Regionen der Kopfkapsel praktisch identisch sind, sollten sich weitere Bemühungen auf den Vergleich höherer Taxa konzentrieren. Allerdings scheint es zwischen den großen Acalles-Arten von den Kanarischen Inseln (Acalles aeonii, Acalles argillosus, Acalles grancanariensis, Acalles sonchi u.a..) und z.B. den Dichromacalles- und Kyklioacalles-Arten vom europäischen Festland nur wenige Unterschiede in der Anzahl und der grundsätzlichen Positionierung der Borsten zu geben. [Bayer & Stüben 2000]
4. Gibt es dafür eine Erklärung? - Die endophytische Lebensweise scheint bei den beinlosen Larven insgesamt zu einer starken Reduzierung aller äußeren Merkmale wie Borstenbüschel oder Sensillen-Anhäufungen geführt zu haben. Wenige cuticulare Mechanorezeptoren reichen offensichtlich aus, um den Larven eine erfolgreiche Entwicklung zu ermöglichen. Kleinere Störungen und „Verletzungen“, die vor allem nach den Häutungen auftreten, scheint die Larve hingegen unbeschadet zu überstehen. Obwohl es daher zwischen den Imagines der Arten innerhalb einer Gattungen erhebliche Merkmalsunterschiede bei den Cryptorhynchinae gibt, spielen offensichtlich – z.B. bei fehlendem Konkurrenzdruck - in der präimaginalen Entwicklung diese Merkmalsausdifferenzierungen keine Rolle. In ihrer endophytischen Lebensweise sind Cryptorhynchinae-Larven offensichtlich „evolutive Minimalisten“!
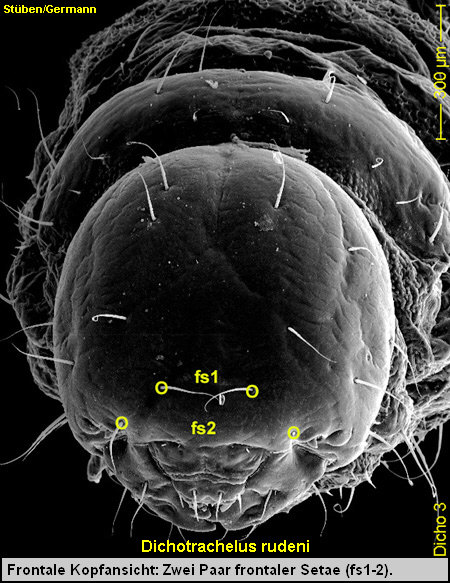
5. Ausdrücklich möchte ich an dieser Stelle auf die jüngste Arbeit von Christoph Germann zu Dichotrachelus rudeni Stierlin 1853 [Germann 2004a, in print] verweisen, in der er zum ersten Mal die Larven dieser Art beschreibt. Sowohl die Lage als auch die Anzahl der Borsten der Kopfkapsel unterscheiden sich erheblich von der Lage und Anzahl der Borsten der Arten der Cryptorhynchinae; hier der Kopfkapsel von Acalles poneli ! So liegen z.B. die frontalen Setae (fs1-2) bei der Larve von Dichotrachelus rudeni in nur 2 Paaren vor. [Fig. Lrud4] Bei den Larven der Cryptorhynchinae sind stets 5 Paare frontaler Setae vorhanden! [Fig. LPon4]
Mein ganz besonderer Dank gilt Christoph Germann (CH-Bern), der mich zum ersten Mal auf einer Kanaren-Reise im Winter 2003/2004 begleitet hat und bei der Suche nach Acalles poneli auf Tenerife einen außergewöhnlichen „Spürsinn“ entwickelte. Wie immer darf ich mich auch bei Peter Sprick (D-Hannover) bedanken, der schon über Jahre hinweg aufmerksam die Diskussion über die so spannende „Dechiffrierung“ der komplexen Biologie der Acalles-Arten begleitet und bereichert. Erneut möchte ich mich auch bei Karin Ulmenkürthen für die Möglichkeit am Bonner Alexander Koenig Museum Rasterelektronenmikroskop-Aufnahmen anfertigen zu dürfen herzlich bedanken.
Bayer,
Chr. & Stüben, P.E. (2000): Vergleichende
Untersuchungen an Larven aus der Acalles-Verwandtschaft. -
Cryptorhynchinae-Studie 14 - in: Stüben, P.E. (2000b), SNUDEBILLER 1 (CD
ROM): 170-181, Mönchengladbach.
Germann Ch. (2004): Entdeckung der potentiellen Wirtspflanze von Acalles anagaensis Stüben
2000 auf Tenerife (España, Islas Canarias) (Coleoptera: Curculionidae:
Cryptorhynchinae) - Weevil News: http://www.curci.de/Inhalt.html,
No. 20, CURCULIO-Institute: Mönchengladbach. (ISSN 1615-3472).
Germann Ch. (2004, in print): Beitrag
zur Biologie von Dichotrachelus rudeni Stierlin, 1853 und Beschreibung
der Larve (Coleoptera: Curculionidae, Cyclominae: Dichotrachelini).
- SNUDEBILLER 5, Studies on taxonomy, biology and ecology of Curculionoidea, Mönchengladbach:
CURCULIO-Institute.
Hegnauer, R. (1966): Chemotaxonomie
der Pflanzen 4. Dicotyledoneae: Daphniphyllaceae - Lythraceae. Basel,
Stuttgart, 551 pp.
Hegnauer, R. (1989): Chemotaxonomie der Pflanzen 8. Nachträge zu
Band 3 und 4 (Acanthaceae bis Lythraceae). Basel, Boston, Berlin, 718 pp.
Kippenberg H. (1983): 25. Unterfamilie Cryptorhynchinae. - Freude,
H.; Harde, K.W. & Lohse, G.A. (Hg.): Die Käfer Mitteleuropas, Bd. 11: 342
S., Krefeld.
Koch K. (1992): Die Käfer Mitteleuropas, Ökologie, Bd. 3, Krefeld, 389
Seiten.
Sprick, P. & Stüben, P.E. (2000): Ökologie der kanarischen
Cryptorhynchinae außerhalb des Laurisilva. - Cryptorhynchinae-Studie 11 - in:
Stüben, P.E. (2000b), SNUDEBILLER 1 (CD ROM): 318-341, Mönchengladbach.
Stüben, P.E. (2000b): (Ed.), Die Cryptorhynchinae der Kanarischen
Inseln. Systematik, Faunistik, Ökologie und Biologie. - SNUDEBILLER 1
(CD ROM); mit 910 Farbfotos, 266 REM-Aufnahmen, 118 Verbreitungskarten, 18 Ton-
u. 1 Video-Aufnahme, 1. Edition,
CURCULIO-Institute, D-Mönchengladbach: 413 pp. (662MB).
Stüben, P.E. (2000e): Die Arten des Genus Acalles von den
Kanarischen Inseln. - Cryptorhynchinae-Studie 5 - in: Stüben, P.E. (2000b),
SNUDEBILLER 1 (CD ROM): 22-98, Mönchengladbach.
Stüben, P.E. (2000k): Phylogenie der endemischen Taxa des Genus Acalles
von den Kanarischen Inseln. - Cryptorhynchinae-Studie 12 - in: Stüben, P.E.
(2000b), SNUDEBILLER 1 (CD ROM): 287-292, Mönchengladbach.
Stüben P.E. (2003d): Breeding of Kyklioacalles euphorbiophilus Stüben
2003 (Coleoptera: Curculionidae: Cryptorhynchinae) - Weevil News: http://www.curci.de/Inhalt.html,
No. 15: 6 pp., CURCULIO-Institute: Mönchengladbach. (ISSN 1615-3472).
Stüben P.E. (2004a): Zucht
von Calacalles droueti (Crotch 1867) von den Azoren (Coleoptera:
Curculionidae: Cryptorhynchinae) - Weevil News: http://www.curci.de/Inhalt.html,
No. 18: 6 pp., CURCULIO-Institute: Mönchengladbach. (ISSN
1615-3472) and - COLEO: http://coleo.de/2004/Calacalles/Calacalles.html,
Nr. 5: S. 1-5,
(ISSN 1616-3281).
Stüben, P.E. (2004b, in print): Revision
der Kyklioacalles teter-barbarus Gruppe - Anmerkungen zur Biologie und
evolutiven Adaptation der neuen Arten (Coleoptera: Curculionidae:
Cryptorhynchinae) - SNUDEBILLER 5, Studies on taxonomy, biology and ecology of
Curculionoidea, Mönchengladbach: CURCULIO-Institute.
Stüben, P.E. (2004d, in print): Die Cryptorhynchinae der Azoren (Coleoptera:
Curculionidae) - SNUDEBILLER 5, Studies on taxonomy,
biology and ecology of Curculionoidea, Mönchengladbach: CURCULIO-Institute.
Adresse
des Autors
Dr. Peter E. Stüben
CURCULIO-Institut
Hauweg 62, D- 41066 Mönchengladbach, Germany
E-Mail: P.Stueben@t-online.de
Internet: www.PeterStueben.de
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}