|
Weevil News |
No.
15 |
6
pp. |
13th
November 2003 |
ISSN
1615-3472 |
|
|
Stüben
P.E. (2003):
Breeding of Kyklioacalles euphorbiophilus Stüben 2003 (Coleoptera:
Curculionidae: Cryptorhynchinae)-
Weevil News: http://www.curci.de/Inhalt.html,
No. 15: 6 pp.,
CURCULIO-Institute: Mönchengladbach. (ISSN 1615-3472). |
|||||
Breeding
of Kyklioacalles euphorbiophilus Stüben
2003
(Coleoptera:
Curculionidae: Cryptorhynchinae)
by
Peter Stüben, Mönchengladbach
with
20 figures and 1 video [Med.Eup]
Abstract
Kyklioacalles
euphorbiophilus Stüben
2003 from High Atlas, Morocco, is bred from dying parts of its host-plant, Euphorbia
nicaeensis All. Taking as a starting-point habitat requirements at the
‘locus typicus’, the biotic and abiotic environmental conditions and methods
of this successful breeding are presented here. All phases of development and
breeding are demonstrated by 20 figures and 1 video sequence. Finally,
recommendations for further breeding of flightless Cryptorhynchinae are given.
Keywords
Coleoptera,
Curculionidae, Cryptorhynchinae, Kyklioacalles euphorbiophilus, breeding, host-plant, Euphorbia
nicaeensis, Morocco.
Introduction
During the work on my first
description of Kyklioacalles
euphorbiophilus in the beginning of 2003 [Stüben 2003a: 134-138], I was
successful in breeding this endemic species from the Moroccan High Atlas.
Kyklioacalles
euphorbiophilus [Fig. 175.1] is closely related to Kyklioacalles
apogeus (Peyerimhoff 1925) [Fig.
176.1]
[Fig. 178.6] from High Atlas near Tachedirt
(‘Tizi-n-Tachedirt’, 3230 m), fallen into oblivion since then. The ‘locus
typicus’ of Kyklioacalles
euphorbiophilus - separated by large mountain massifs - is situated
about 60 km south-westerly on the northern face of the ‘Tizi-n-Test’ (2000
m) [Fig.
175.20]. The species is strictly bound to Euphorbia
nicaeensis
All. and lives on steep slopes of maximum 1-2 hectars. This extremely unusual
situation just even produced the assumption that this big Kyklioacalles
species
from
the
Kyklioacalles teter-/barbarus-group could be a new species (see
'bionomy'). Reproductive isolation after separation by huge mountain massifs and
subsequent specialisation to a - here also very isolated growing -
host-plant really seem to be important factors in species separation.
Bionomy
A
lot of specimens of the new species were sieved from a very wet slope on the
north side of Tizi-n-Test in the Moroccan High Atlas and from detritus and roots
of Euphorbia nicaeensis All. Strong
specimens of this Euphorbiaceae occurring totally isolated here grew between
broom shrubs and were often damaged by the skidding scree - to sprout again from
the very lignified parts of the trunk (shoots). [Fig.
175.20] To compare with Euphorbia, the neighbouring broom shrubs were also sieved
intensively. But not a single specimen could be obtained from the massive,
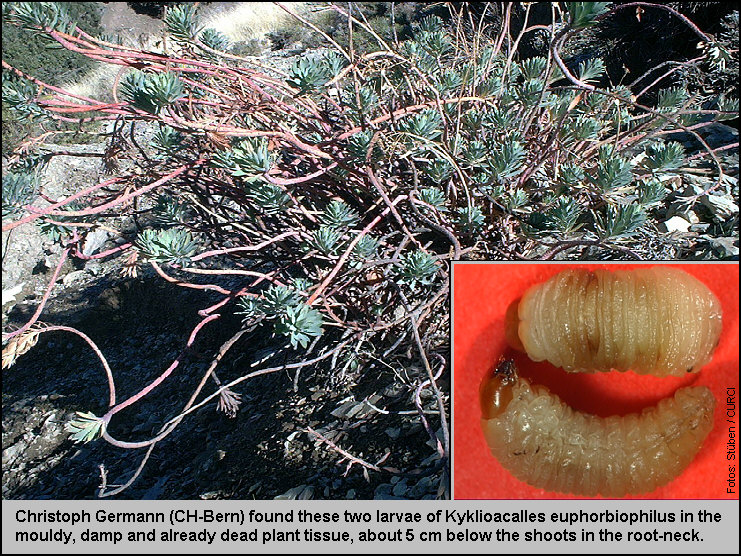
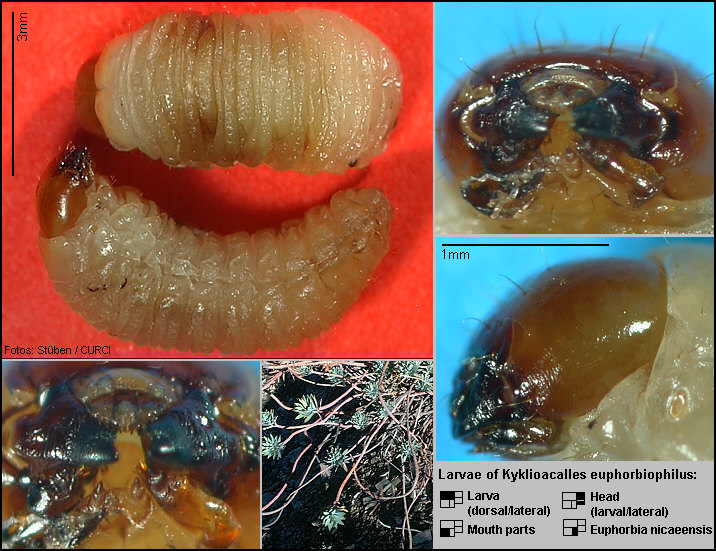
damp-mouldy detritus! On the other hand, Christoph Germann (Bern) detected two
larvae of the new species in a strong, dead specimen of Euphorbia nicaeensis. [Fig. 175.21]
For this reason, Euphorbia nicaeensis
was teared open from upside down to the root. He found the larvae in the mouldy,
damp plant tissue, about 5 cm below the shoots in the root-neck. [Fig.
175.22] These larvae must be ‘stragglers’, because larvae of all
known Kyklioacalles species normally
develop during midsummer (in the tissue of dying plants). In the partly dead
root-neck of other plants further feeding-caverns of the larvae were found.
Euphorbia
nicaeensis shows
a tendency to form biological races. The
complex has a Mediterranean-south-Siberian distribution (steppicol); but Euphorbia nicaeensis sensu strictu has a western and central
Mediterranean distribution. A population from Toscana with a creeping habit (7 -
15 cm) and with 3 - 8 umbel rays was described as var. prostrata Caruel, an Apulian with hairy capsule as var. japygica
(Ten.) Arcang. [Pignatti 1982]. Therefore, in the Mediterranean area it is a
wide-spread species, often showing local forms or it is a species complex. The
completely isolated occurrence of this member of the Spurge family 2000 m above
sea level - we didn’t find this plant at no other locality of the High Atlas!
- nurtures the suspicion that the little isolated population of Tizi-n-Test
could be a separate form of E. nicaeensis.
On no other Euphorbiaceae we found Kyklioacalles
species in the Rif Mountains, Middle Atlas, High Atlas or Anti-Atlas. So, I
assume that Kyklioacalles euphorbiophilus
in an endemic species of the central mountains of High Atlas (if not even of
Tizi-n-Test).
Methods
These very
detailed observations in the Moroccan High Atlas and especially the fact, that Kyklioacalles
euphorbiophilus develops inside the root-neck of dying Euphorbia
nicaeensis-plants, encouraged me to breed a Cryptorhynchinae (here: a Kyklioacalles
species) for the first time at all. For this purpose I brought a small
fresh, uncolonized specimen of Euphorbia
nicaeensis to Germany. In a customary flat clay-flowerpot with a diameter of
18 cm and a height of 9 cm, the plant was put into a mixture of sand, a little
bit of loam and small bloated clay-balls, as it is used for hydroponics, on
January 6, 2003. The bloated clay-balls are used as drainage, to protect from
damp and early growth of fungi. But transport and not very careful treatment of
the plant in the insectarium of the CURCULIO-Institute caused a gradually dying
of the outer branches. A reaction of the plant was to produce
‘trouble-shoots’. [Fig. Z2]. This was even intended, because at ‘locus
typicus’ mainly plants are colonized being damaged by the skidding scree, to
sprout again from the lignified segments of the trunk.
On
January 8, 2003, I put 6 females and 5 males in the glass-insectarium of the
size: 30 cm x 30 cm x 40 cm (length x width x height). There were also 4 cm of
sand around the pot, pressed down slightly and watered regularly. [Fig.
Z2] The plant itself was still treated ‘uncarefully’, only sprayed in
two-day intervals and watered weekly and sparsely. The animals oriented towards
the plant immediately after they were put in; at night they climbed the branches
of the plant and during the day they retreated to the clay-balls deeply between
the roots. In the evening twilight the weevils climbed the stems and copulated
here. They were observed feeding from dead parts of the plant
(‘rasp-feeding’) [Fig. 175.23]. The plant was
taken out by night together with the pot to document the events with a
high-resolution digital camera. [Fig.
175.23]
[Med.Eup].
Breeding
In
March, the plant tried to produce shoots for a last time, before drying up [Fig.
Z2]. The first 4 larvae could be taken from the completely dried
root-neck on May 21, 2003. [Fig. Z1]. They had
reached the last larval stage. On June 10, 2003, I found the two first pupae in
their pupal chambers, filled with used rasp-shavings. Astonishingly, the
root-neck and the trunk, projecting about 5 cm and containing the larvae,
remained relatively damp [Fig. Z3]. Only
occasionally the plant was watered and slightly sprayed in its dried state. The
temperature in the insectarium was 21 °C until end of April, and afterwards
about 26 °C. The larvae were found in the primary root, the subterranean
root-neck, in the trunk and in the lower small forks of the branches of Euphorbia
nicaeensis; but not in the dried twigs.
On
June 23, 2003, the emerging of the first immature specimen could be photographed
[Fig. Z4]. The egg-shaped pupal chamber was 8 mm
long and 4.5 mm wide. To film the emerging from the pupa, the pupal chamber was
partly opened on one side [Fig. Z5]. On the same
day, further photographs were taken of an already far developed pupa. The pupa
was put into a small Eppendorf-vessel and covered with dead, small-rasped
substratum of the Euphorbiaceae. In the apical part of the Eppendorf-vessel, the
pupa made a new pupal chamber by violent movement [Fig.
Z6].
On
June 26, 2003, further photographs were taken from the same pupa; the dark eyes
are clearly to be recognized already now [Fig. Z7].
On July 1 or 2, 2003, the first animal must have emerged, because on July 3,
2003, I found a very immature (but no more white-colored) imago.
On
June 30, 2003, the dismantling of the trunk segment was continued. It was found:
1 still white imago [Fig. Z8]; it was put in a small Eppendorf-vessel. 4 days later, it was completely hardened,
1
completely hardened imago in its pupal chamber,
1
pupa; the imago emerged on July 2, 2003; but it stayed in its pupal chamber
until mortification on July 6, 2003.
1
larva of the last larval stage [Fig. Z11] and
further larvae (of all larval stages) and pupae in the more dry, upper part of the lignified trunk; right below the broken twigs long time ago. The root- and twigless trunk-segment (about 3 x 5 cm) was photographed [Fig. Z9] and put back to the original place. It seems to be possible, that such small trunks are transported over long distances by wind or water and that they are responsible for the main spreading of the species: Larvae and pupae and (completely hardened) adults can obviously spend weeks or even months inside these small trunks. [Fig. Z10] They are outstandingly well protected and only wait for the total breakdown of the small trunks, instead of being partly decomposed by the activity of the larvae. They don’t show any own efforts to free themselves! Some larvae, pupae and a still white imago were put in Eppendorf-vessels for further development. They could be bred to complete imagos under constant observation.
On
July 6, 2003, the roots already separated from the above-ground trunk segment
were digged out; then, from the central,
damp and totally rasped
root-area 3 pupae, 1 big and 1 small larvae could be taken. [Fig.
Z12] Then, the trunk segment still containing pupae and larvae was
preserved. [Fig. Z9] Not until July 9, 2003, the
last larvae were taken from the trunk segment, deadened and boiled with water
and finally preserved in alcohol.
(It
is not necessary to tell, that a continuation of breeding was not intended,
because the host-plant Euphorbia
nicaeensis (from the Moroccan High Atlas) didn’t stand at my disposal).
Discussion
and Preview
From
this first successful breeding (as I know) of a Cryptorhychinae unable to fly, a
member of the genus Kyklioacalles,
first clues for a more standardized breeding could be obtained.
1.
It seems that only plants during the process of dying make
possible this success. Dead wood possibly still contains larvae and pupae, but
the females don’t use it for laying eggs, in my mind. All experiences
from my studies on the Macaronesian islands clearly show, that Cryptorhynchinae
belong to the very first colonizers of such plants, that are in the process of
dying (f.e. introduced by wind-break). [Stüben 2000b][Stüben 2000j][Stüben
2000][Sprick & Stüben 2000]; Cossoninae are following much later (f.e. Mesites,
Cossonus, Phloeophagus). Therefore, stressed plants or even fresh plant-parts
should be sufficient, to start a successful breeding. It is important, that the
supply of toxic plant compounds - as in Euphorbiaceae - was already stopped to a
great extent!
2.
Larvae of Cryptorhynchinae seem to be extremely enduring and
subsist on very little. They survived even a drying for weeks of the upper
trunk-segments and forks of the twigs in March 2003 without any damage. (I was
on an excursion on this point in time). For that reason, the whole substratum
for development should be kept relatively dry, and it should be drained
sufficiently.
3.
After
emerging
from the pupa, the adult weevils still stay for weeks, if not for months, in
their pupal chambers. Not before the first precipitation in autumn and/or winter
and softening of plant tissue, the animals free themselves from their
substratum, in which they have developed. [Stüben 2000e] An active spreading
seems to be very improbable for these flightless Cryptorhynchinae, and it might
occur only very rarely. Passive transport of the died and completely dried
developmental substratum by wind or water should play a much more crucial role
for the colonization of new habitats. (The suspicion that lignified flotsam
could also play an important role for the (first) colonization of islands,
doesn’t seem to be false). Therefore, twigs, branches and parts of the roots
should be cut open regularly, to be able to document the phase of metamorphosis!
At
this place I am unfortunately not allowed to describe the eggs, larvae and pupae
of Kyklioacalles euphorbiophilus and
to designate holo- and paratypes, according to the ‘International Code of
Zoological Nomenclature’ [ICZN: 2000: § 9.8.]; see [Bayer & Stüben 2000]. Unfortunately, contributions with
nomenclatural actions still may not be published in the World Wide Web - as is fixed by the commission. Therefore, I have to
turn attention of the readers to our next SNUDEBILLER 5 / 2004: Here will be
catched up, what could have been better documented at this place and in this
context.
Acknowledgements
My special thanks
is dedicated to Christoph Germann (Bern/Switzerland). We discovered the new
species Kyklioacalles euphorbiophilus
at Euphorbia nicaeensis in the High
Atlas on one of our numerous common journeys to Morocco. My thanks is also
expressed to Peter Sprick (Germany/Hannover), who translated this contribution
into English.
References
Bayer, Chr. & Stüben, P.E. (2000): Vergleichende Untersuchungen an
Larven aus der Acalles-Verwandtschaft.
- Cryptorhynchinae-Studie 14 - in: Stüben, P.E. (2000b), SNUDEBILLER 1 (CD
ROM), CURCULIO-Institute, D-Mönchengladbach: 170-181
Pignatti,
S.
(1982): Flora d’Italia. Vol. secondo. Bologna, 732 pp.
Sprick,
P. & Stüben, P.E. (2000):
Ökologie der kanarischen Cryptorhynchinae außerhalb des Laurisilva. -
Cryptorhynchinae-Studie 11 - in: Stüben, P.E. (2000b), SNUDEBILLER 1
(CD ROM), CURCULIO-Institute, D-Mönchengladbach: 318-341
Stüben,
P.E. (2000b):
(Ed.), Die Cryptorhynchinae der Kanarischen Inseln. Systematik, Faunistik, Ökologie
und Biologie. - SNUDEBILLER 1 (CD
ROM); mit 910 Farbfotos, 266 REM-Aufnahmen, 118 Verbreitungskarten, 18 Ton- u. 1
Video-Aufnahme, 1. Edition,
CURCULIO-Institute, D-Mönchengladbach: 413 pp.
Stüben,
P.E. (2000e):
Die Arten des Genus Acalles von den
Kanarischen Inseln. - Cryptorhynchinae-Studie 5 - in: Stüben, P.E. (2000b),
SNUDEBILLER 1 (CD ROM),
CURCULIO-Institute, D-Mönchengladbach: 22-98
Stüben,
P.E. (2000j):
Biogeographie und Evolution der kanarischen Cryptorhynchinae. -
Cryptorhynchinae-Studie 10 - in: Stüben, P.E. (2000b), SNUDEBILLER 1
(CD ROM), CURCULIO-Institute, D-Mönchengladbach: 293-306
Stüben,
P.E. (2002):
Die Cryptorhynchinae von den Inseln
Madeiras und Salvagens. Taxonomie, Bionomie, Biogeographie und Evolution.
(Coleoptera: Curculionidae). - SNUDEBILLER 3,
Studies on taxonomy, biology and ecology of Curculionoidea, Mönchengladbach:
CURCULIO-Institute, 88-195.
Stüben,
P. E. (2003a): Revision
des Genus Kyklioacalles und
Beschreibung der Untergattung Palaeoacalles
subg. n. unter Heranziehung phylogenetischer, morphogenetischer und
biogeographischer Aspekte (Curculionidae: Cryptorhynchinae). - SNUDEBILLER 4,
Studies on taxonomy, biology and ecology of Curculionoidea, Mönchengladbach:
CURCULIO-Institute: 116 - 166.
Address:
Dr.
Peter E. Stüben
CURCULIO-Institute
Hauweg
62
D-41066 Mönchengladbach
E-Mail:
P.Stueben@t-online.de
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}